서론:
현재 존재하는 단백질들은 수십억 년에 걸친 자연 진화를 통해 현재의 형태로 발전해 왔습니다. 이 과정에서 무작위적인 돌연변이들이 발생하고, 생물학적 선택을 통해 단백질들은 그들의 서열, 구조, 기능에 따라 걸러졌습니다. 이러한 과정에서 형성된 패턴들은 생물학적 진화의 숨겨진 변수들이 오랜 시간 동안 단백질의 변형을 어떻게 이끌어왔는지에 대한 정보를 담고 있습니다.
지구상의 생물 다양성을 조사하는 유전자 시퀀싱 연구들은 이미 단백질 서열과 구조에 대한 방대한 양의 데이터를 축적해 왔으며, 이러한 데이터는 생명체 간의 변이 패턴을 조명하는 데 기여하고 있습니다. 이러한 데이터를 통해 서열 뒤에 숨어있는 단백질 생물학의 근본적인 언어가 존재한다는 합의가 형성되고 있으며, 이를 언어 모델을 사용해 이해할 수 있다는 것이 이 논문의 핵심 주장입니다.
단백질 서열에 대한 언어 모델의 연구:
최근 들어 여러 연구에서 단백질 서열에 대한 언어 모델들이 개발되고 평가되고 있습니다. 이 언어 모델들은 단백질의 생물학적 구조와 기능을 반영하는 표현을 학습할 수 있으며, 이러한 학습은 단백질의 특성에 대한 명시적인 감독 없이도 가능하다는 것이 입증되었습니다. 이 모델들은 스케일이 커질수록 성능이 향상되며, 이러한 스케일 확장은 컴퓨팅 능력, 파라미터, 데이터 측면에서 발전의 전선에 서 있습니다.
ESM3 소개:
ESM3는 서열, 구조, 기능을 종합적으로 고려할 수 있는 최첨단 다중모달 생성 모델로, 3차원 원자 구조를 복잡한 건축이나 3차원 공간 내의 확산을 사용하지 않고, 이 구조를 불연속 토큰으로 변환해 학습합니다. 이 방식은 확장 가능하며, 다양한 입력 조합에 맞춰 새로운 단백질을 생성하는 데 적합합니다. ESM3는 대규모 모델일수록 더 나은 성능을 보이며, 모든 스케일의 모델이 조정에 더 민감하게 반응합니다.
결론적으로, ESM3는 수십억 개의 파라미터를 바탕으로 단백질 서열, 구조, 기능을 성공적으로 학습하고, 단백질 생성을 제어할 수 있는 가능성을 보여주는 모델입니다.
이 서론에서는 단백질 진화의 복잡한 메커니즘을 모방하여 기능성 단백질을 새롭게 설계할 수 있는 가능성을 제시하고 있으며, 언어 모델을 통해 단백질의 변이 패턴을 학습할 수 있는 방법론을 설명하고 있습니다.
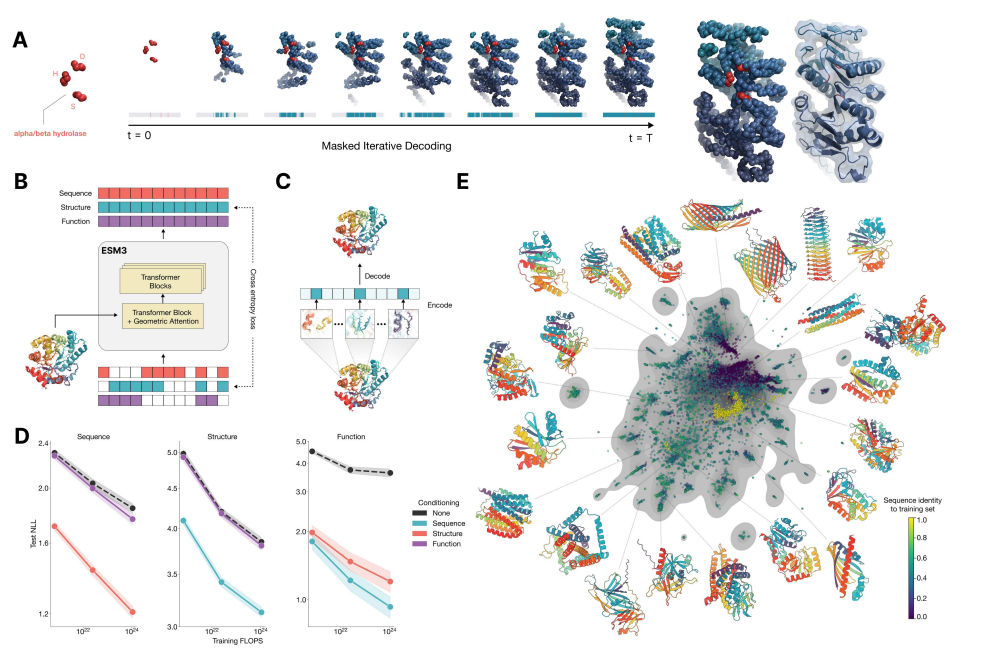
ESM3 모델 설명:
ESM3는 서열(단백질의 아미노산 서열), 구조(단백질의 3차원 구조), 기능(단백질이 수행하는 역할)을 모두 고려하는 다중모달(다양한 데이터를 처리할 수 있는) 모델입니다. 이 모델은 인공지능의 한 종류인 트랜스포머(Transformer) 모델을 기반으로 하고 있으며, 이는 언어를 이해하거나 번역하는 데에 주로 사용되던 모델입니다. 이 트랜스포머는 한 문장에서 단어들 간의 관계를 이해하듯이, 단백질 서열 안의 아미노산들 간의 관계도 학습할 수 있습니다.
비유를 사용한 설명:
트랜스포머 모델을 이해하기 위해, '퍼즐 맞추기'에 비유할 수 있습니다. 단백질 서열은 퍼즐 조각이고, 그 조각들이 어떻게 결합하여 완전한 그림(단백질의 기능과 구조)을 이루는지를 모델이 학습합니다. 예를 들어, 단백질 서열이 제공되면 ESM3 모델은 이 조각들이 서로 어떻게 연결되어야 할지를 예측하고, 그 결과로 단백질이 어떻게 접히고 기능할지 예측할 수 있습니다.
1. 서열 예측:
모델은 먼저 단백질의 아미노산 서열을 입력받습니다. 이는 마치 퍼즐 조각들을 한 줄로 늘어놓은 것과 같습니다. 각 조각(아미노산)은 특정한 위치에 올바르게 놓여야 전체 퍼즐이 완성됩니다. 이때 ESM3는 각각의 퍼즐 조각이 어디에 위치해야 하는지, 그리고 각 조각이 서로 어떤 관계를 가지고 있는지를 예측합니다. 이를 통해 모델은 다음 아미노산이 무엇이 되어야 하는지를 알아냅니다.
2. 구조 예측:
단백질의 3차원 구조를 이해하는 것은 마치 퍼즐 조각들이 2차원이 아닌 3차원에서 맞물리는 것과 같습니다. 이 모델은 아미노산 서열이 단백질의 3차원 구조로 어떻게 접히는지를 예측합니다. 여기서 중요한 개념은 입체 구조입니다. 퍼즐 조각들이 서로 연결될 때, 어떤 조각은 위로, 어떤 조각은 아래로, 혹은 옆으로 꼬일 수 있습니다. 이러한 공간적 관계를 예측하는 것이 바로 구조 예측의 핵심입니다.
이 구조 예측에서 중요한 요소는 구조 토큰화(tokenization)입니다. 마치 문장을 단어 단위로 나누듯이, 단백질의 3차원 구조를 '토큰'이라는 작은 단위로 나눕니다. 각 아미노산은 자신만의 구조를 가지는데, 이 구조는 단순히 그 위치에 해당하는 좌표와 각도로 정의됩니다. 이를 통해 ESM3는 각각의 아미노산이 주변 환경과 어떻게 상호작용하는지를 더 쉽게 이해할 수 있게 됩니다.
3. 트랜스포머 블록:
ESM3의 핵심은 트랜스포머 블록입니다. 트랜스포머는 여러 층(layer)으로 이루어져 있고, 각 층에서는 입력된 데이터(서열, 구조, 기능)들이 서로 연결되고 상호작용하는 방식을 학습합니다. 예를 들어, 트랜스포머는 한 아미노산이 어떤 다른 아미노산들과 연결되어 있는지, 그리고 그 관계가 어떻게 단백질의 기능에 영향을 미치는지를 파악합니다. 이를 위해 자기 주의 메커니즘(self-attention mechanism)이라는 중요한 개념이 사용됩니다.
4. 자기 주의 메커니즘 비유:
이 메커니즘은 마치 교실에서 선생님이 학생들을 바라보며 누가 중요한 질문을 하고 있는지, 혹은 누가 집중해야 할지 파악하는 것과 비슷합니다. 트랜스포머의 각 층에서는 모든 아미노산이 서로를 '바라보고'(주의를 기울이며) 이 중에서 가장 중요한 정보를 선택합니다. 이 과정을 통해 모델은 단백질 내에서 어떤 아미노산들이 중요한 역할을 하는지를 배웁니다.
5. 예측의 확률적 성격:
모델은 단백질을 생성하거나 예측할 때 확률적으로 접근합니다. 이는 마치 퍼즐 조각이 여러 위치에 놓일 수 있는 가능성을 고려하는 것과 같습니다. ESM3는 각 아미노산 서열의 각 위치에서 가장 적합한 조각을 찾는 확률을 계산하여 단백질 구조를 완성합니다. 이러한 확률적 접근을 통해 모델은 더 정확하게 예측을 수행할 수 있습니다.
6. 모델의 학습:
ESM3는 방대한 데이터에서 학습을 합니다. 마치 퍼즐을 풀 때 다양한 그림을 통해 학습하듯이, 이 모델은 수많은 단백질 데이터를 통해 서열, 구조, 기능 간의 관계를 이해하게 됩니다. 이 과정에서 손실 함수(loss function)를 사용하여 예측이 얼마나 정확한지를 평가하고, 모델이 더 나은 결과를 내도록 지속적으로 학습합니다.
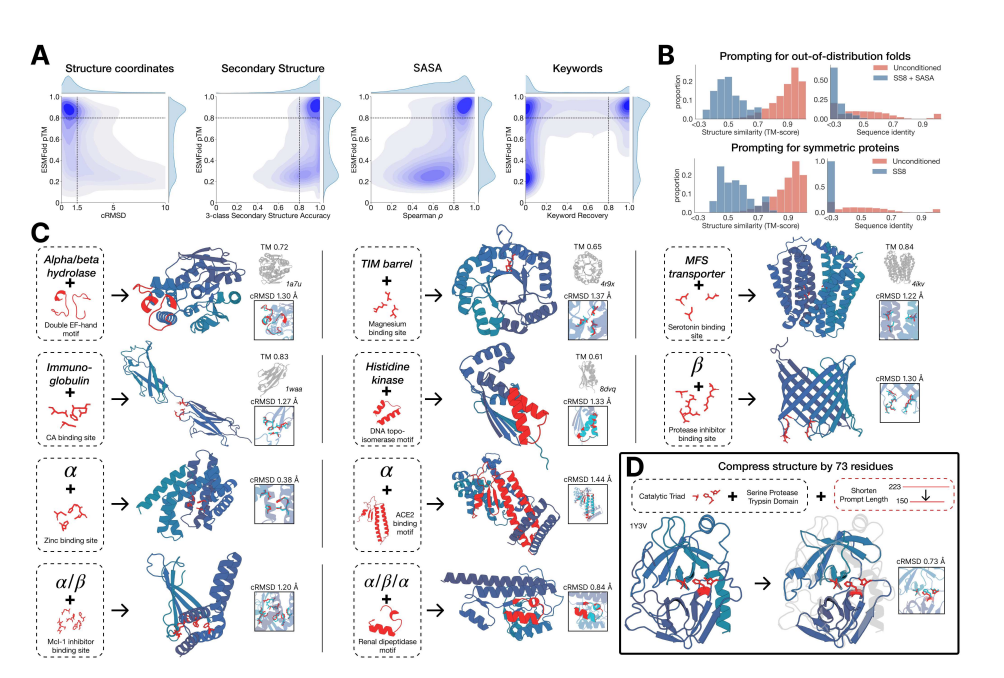
ESM3의 학습 과정과 토큰화
1. 구조 토큰화 (Structure Tokenization):
ESM3는 단백질의 3차원 구조를 학습할 때, 그 구조를 "토큰(token)"이라는 단위로 나눕니다. 비유하자면, 마치 복잡한 퍼즐을 완성할 때 각 조각을 세부적으로 분해하여 더 작은 부분으로 나누는 것과 비슷합니다. 이 과정에서 각 아미노산의 3차원 구조는 주변 아미노산들과의 상대적 위치와 방향을 포함합니다. 이를 통해 모델은 단백질의 복잡한 공간적 배열을 더 쉽게 이해하고 예측할 수 있습니다.
구체적으로, 각 아미노산의 위치는 프레임(frame)이라는 개념으로 정의됩니다. 이 프레임은 그 아미노산의 위치와 방향을 나타내며, 다른 아미노산들과의 상대적 거리와 각도에 따라 달라집니다. 여기서 중요한 것은 SE(3) 변환이라는 수학적 개념입니다. 이를 쉽게 설명하자면, 하나의 아미노산을 다른 아미노산에 대해 회전시키거나 이동시키는 방법을 수학적으로 나타내는 방식입니다.
비유:
이를 더 이해하기 쉽게 설명하자면, 각 아미노산은 마치 '나침반' 같은 역할을 합니다. 이 나침반은 단백질 구조 내에서 다른 아미노산들이 어느 방향으로, 얼마나 떨어져 있는지를 알려줍니다. ESM3는 이러한 나침반을 사용하여 각 아미노산이 전체 구조에서 어떻게 배치되어 있는지를 계산합니다.
2. 기하학적 주의 메커니즘 (Geometric Attention Mechanism):
ESM3는 단백질의 3차원 구조를 예측할 때, 단순히 서열상의 정보뿐만 아니라 기하학적 정보(즉, 아미노산들 간의 실제 거리와 방향)를 고려합니다. 이를 위해 기하학적 자기 주의 메커니즘을 사용합니다. 이 메커니즘은 모든 아미노산들이 서로의 3차원 구조를 고려하여 상호작용하는 방식을 학습하는 과정입니다.
비유하자면, 마치 우리가 무언가를 조립할 때 각각의 부품들이 서로 어떤 각도와 방향으로 연결되어야 하는지 고려하는 것과 같습니다. 예를 들어, 자동차를 조립할 때 바퀴와 차체가 정확한 위치와 방향으로 조립되지 않으면 제대로 작동하지 않는 것처럼, 단백질도 아미노산들이 정확한 위치에 있어야 제대로 기능합니다. 기하학적 자기 주의 메커니즘은 이러한 관계를 학습하여 단백질의 3차원 구조를 예측합니다.
3. 단백질 구조 재구성:
ESM3는 학습을 통해 새로운 단백질 구조를 생성할 수 있습니다. 이 과정에서 VQ-VAE(벡터 양자화 변이 오토인코더)라는 기법을 사용합니다. 이는 복잡한 정보를 간단한 코드(토큰)로 압축하여 저장하고, 나중에 다시 복원하는 방식입니다. 예를 들어, ESM3는 단백질의 복잡한 구조를 작은 토큰들로 나눈 후, 이를 조합하여 새로운 구조를 만들어냅니다.
비유:
이를 마치 '레고 블록'에 비유할 수 있습니다. ESM3는 레고 블록(토큰)을 가지고 다양한 단백질 구조를 만들 수 있습니다. 레고 블록 하나하나는 단순하지만, 그것을 조합하면 복잡한 구조물이 만들어집니다. ESM3가 학습한 것은 이 레고 블록들을 효율적으로 조합하여 새로운 단백질을 만드는 방법입니다.
4. 학습 과정에서의 노이즈 추가 (Adding Noise in Training):
모델이 학습할 때는 노이즈(잡음)를 추가하는 과정이 있습니다. 이는 모델이 주어진 정보에서 누락된 데이터를 채워 넣는 법을 학습하기 위함입니다. 예를 들어, 단백질의 서열이나 구조 중 일부를 가리고, 모델이 나머지 부분을 보고 누락된 부분을 예측하도록 훈련합니다.
비유:
이 과정을 퍼즐 맞추기에 비유하면, 퍼즐의 일부 조각을 가리고 나머지 조각들만 보고 가려진 조각이 어디에 있어야 하는지 추론하는 것과 비슷합니다. ESM3는 이러한 방식으로 단백질 서열이나 구조의 일부가 주어졌을 때, 나머지를 예측할 수 있는 능력을 키웁니다.
5. 요약:
ESM3는 단백질의 서열, 구조, 기능을 종합적으로 이해하고 새로운 단백질을 생성할 수 있는 모델입니다. 이 모델의 핵심은 트랜스포머 아키텍처와 기하학적 자기 주의 메커니즘, 그리고 벡터 양자화 기법을 통해 매우 복잡한 단백질 구조를 효율적으로 학습하고 예측할 수 있다는 것입니다.
계속해서 ESM3의 학습과 예측 과정에 대해 구체적으로 설명하겠습니다. 여기서는 모델이 어떻게 실제 단백질을 생성하거나 예측하는지, 그 메커니즘에 대해 더 깊이 다뤄보겠습니다.
6. 확률적 샘플링 (Probabilistic Sampling):
ESM3는 단백질 서열이나 구조를 예측할 때 확률적으로 접근합니다. 즉, 한 가지 답을 고정적으로 내놓는 것이 아니라, 여러 가능성 중에서 가장 적합한 것을 선택합니다. 이는 확률적 샘플링 기법을 통해 이루어집니다.
비유:
이 과정은 마치 여러 개의 문항 중에서 가장 가능성이 높은 답을 선택하는 과정과 비슷합니다. 시험 문제에서 정답이 여러 개일 수 있고, 각 정답의 가능성에 점수를 매기는 것처럼, ESM3는 각 아미노산 서열의 각 위치에서 가능성이 가장 높은 아미노산을 확률적으로 선택합니다. 이를 통해 다양한 서열을 생성하거나 예측할 수 있습니다.
확률적으로 예측하는 이유는 단백질의 구조나 기능이 매우 복잡하고, 여러 변수가 상호작용하기 때문에 한 가지 결정적 답이 아닌, 여러 가능성의 조합을 고려할 필요가 있기 때문입니다.
7. 토큰 샘플링 과정 (Token Sampling Process):
ESM3는 여러 단계의 예측 과정을 통해 단백질 서열을 생성합니다. 각 단계에서 모델은 단백질의 서열이나 구조의 일부를 마스크(mask)한 후, 나머지 정보를 바탕으로 마스크된 부분을 예측합니다. 이때 예측은 단 한 번에 모든 위치를 채우는 것이 아니라, 일부 위치를 먼저 채우고, 이를 기반으로 다시 나머지 위치를 채우는 과정을 반복합니다.
비유:
이를 퍼즐 맞추기로 비유하자면, 처음에는 가장자리 부분부터 맞추기 시작하고, 점차 중앙 부분을 채워가는 방식입니다. 처음부터 모든 퍼즐 조각을 한 번에 맞추려고 하는 것이 아니라, 각 단계를 거치면서 점점 더 많은 퍼즐 조각들을 맞추어 완성도를 높여가는 방식입니다.
8. 트랜스포머 구조의 특성:
트랜스포머는 여러 층(layer)으로 이루어진 모델입니다. 각 층은 이전 층의 출력(output)을 받아 다음 층에서 더 복잡한 패턴을 학습합니다. 이를 통해 단백질의 서열이나 구조에 대한 더 높은 수준의 정보를 얻을 수 있습니다.
비유:
이 과정을 건물의 층으로 비유할 수 있습니다. 첫 번째 층에서는 단순한 정보를 처리하고, 두 번째 층에서는 그 정보를 바탕으로 더 복잡한 패턴을 찾으며, 마지막 층에서는 전체적인 구조를 완성하는 것입니다. 각 층을 거칠수록 모델은 더 복잡한 관계와 패턴을 이해하게 됩니다.
9. 기하학적 주의 메커니즘을 통한 구조 예측:
ESM3는 단백질의 서열뿐만 아니라, 그 서열이 어떻게 3차원 구조로 접히는지도 예측합니다. 이를 위해 기하학적 주의 메커니즘을 사용하여 아미노산 간의 공간적 관계를 파악합니다. 이 과정에서 각 아미노산의 위치와 방향, 그리고 서로 간의 거리를 고려합니다.
비유:
이 과정을 마치 3D 퍼즐을 맞추는 것에 비유할 수 있습니다. 퍼즐의 각 조각이 단순히 평면 위에 놓이는 것이 아니라, 3차원 공간에서 회전하거나 이동하면서 서로 연결되어야 합니다. ESM3는 이러한 공간적 관계를 이해하고 예측할 수 있습니다.
10. 모델 평가 (Model Evaluation):
모델을 평가하는 데에는 여러 가지 방법이 있습니다. 예를 들어, 단백질 서열이 주어졌을 때 모델이 얼마나 정확하게 그 단백질의 3차원 구조를 예측하는지를 평가할 수 있습니다. 이를 위해 LDDT (Local Distance Difference Test)와 같은 지표를 사용합니다. LDDT는 모델이 예측한 단백질 구조가 실제 구조와 얼마나 일치하는지를 측정하는 지표입니다.
비유:
이를 학생의 시험 성적에 비유할 수 있습니다. 학생이 문제를 얼마나 정확하게 풀었는지를 평가하는 것처럼, LDDT는 모델이 단백질 구조를 얼마나 정확하게 예측했는지를 평가합니다.
11. 생성 모델로서의 ESM3:
ESM3는 단백질을 예측하는 것뿐만 아니라, 아예 새로운 단백질을 생성할 수도 있습니다. 이는 단백질의 서열이나 구조를 전혀 모르는 상태에서도 새로운 단백질을 만들어내는 능력을 의미합니다.
비유:
이를 요리사에 비유할 수 있습니다. 기존의 레시피를 보고 요리를 만드는 것이 아니라, 주어진 재료만으로 새로운 레시피를 만들어서 요리를 완성하는 것과 같습니다. ESM3는 기존의 단백질 데이터에서 학습한 정보를 바탕으로 새로운 단백질을 만들어낼 수 있습니다.
12. ESM3의 창의적 해결책:
ESM3는 단백질을 예측하거나 생성할 때, 생물학적으로 알려지지 않은 새로운 단백질을 만들어낼 수 있습니다. 이는 수백만 년에 걸친 진화를 통해 자연에서 만들어졌을 가능성이 있는 단백질을 시뮬레이션하는 과정으로 볼 수 있습니다.
비유:
이를 창의적인 발명가에 비유할 수 있습니다. 발명가는 기존에 없던 새로운 도구나 기계를 만들어내듯이, ESM3는 기존에 없던 단백질을 만들어내는 창의적인 역할을 합니다. 이러한 과정에서 ESM3는 생물학적 기능을 유지하면서도 매우 다양한 단백질 구조를 생성할 수 있습니다.
13. 생물학적 정렬(Biological Alignment):
모델의 성능을 높이기 위해 ESM3는 학습 중에 특정 기준을 충족하도록 정렬(Alignment) 과정을 거칩니다. 기본적으로 ESM3는 훈련 데이터에 기반해 단백질 서열을 생성하지만, 단백질의 특정 기능을 충족하기 위해 더 세부적인 조건들을 맞춰야 할 때가 있습니다. 이때 모델을 세부적으로 조정하여 생물학적 기능에 더욱 맞는 단백질을 생성할 수 있습니다.
비유:
이 과정을 요리사의 요리법 조정에 비유할 수 있습니다. 요리사가 새로운 요리를 만들었을 때, 고객이 특정 맛을 원하면 그에 맞춰 레시피를 약간 수정해야 할 때가 있습니다. 마찬가지로, ESM3는 특정 단백질 기능이나 구조를 더 잘 맞추기 위해 조정됩니다.
14. 정렬의 목적:
ESM3는 처음에는 무작위로 단백질을 생성하지만, 이를 더 정확하게 생물학적으로 일치시키기 위해 추가적인 피드백 과정을 거칩니다. 이 피드백 과정은 두 가지 목표를 갖습니다:
- 정확성: 모델이 예측한 단백질이 실제로 기능할 수 있도록 정확한 구조를 갖도록 함.
- 창의성: 모델이 기존의 자연에서 존재하지 않는 새로운 단백질을 만들더라도 생물학적으로 기능할 수 있는지 확인.
비유:
이는 마치 제품 디자인에서 '기능성'과 '창의성'을 동시에 고려하는 것과 같습니다. 새로운 디자인은 참신해야 하지만, 그 디자인이 실제로 유용해야 하듯, ESM3도 새로운 단백질을 창의적으로 생성하면서도 그 단백질이 실제로 기능해야 합니다.
15. 정렬 과정에서의 알고리즘:
정렬을 위해 사용되는 알고리즘은 선호 조정(preference tuning)이라는 방식입니다. 이 알고리즘은 모델이 생성한 여러 샘플 중에서 더 나은(좋은) 샘플과 덜 나은(나쁜) 샘플을 비교하여 학습하는 방식입니다. 좋은 샘플을 선호하도록 모델을 학습시키고, 나쁜 샘플은 피하도록 조정하는 것입니다.
비유:
이 과정은 마치 여러 음식의 맛을 비교하는 시식 과정을 떠올릴 수 있습니다. 시식에서 가장 맛있는 음식을 선호하게끔 평가하는 것처럼, ESM3는 가장 적합한 단백질 구조와 기능을 선호하도록 학습됩니다.
16. 선호 조정 알고리즘의 구조:
선호 조정은 다음과 같은 두 가지 단계로 이루어집니다:
1. 좋은 샘플과 나쁜 샘플 비교: 모델은 여러 번의 샘플을 생성하고, 이 중에서 더 나은 결과를 선택합니다. 이때 더 좋은 구조를 가진 단백질이나, 생물학적으로 더 일치하는 단백질이 선택됩니다.
2. 좋은 샘플을 선호하도록 학습: 선택된 좋은 샘플을 기반으로 모델을 다시 학습시켜, 향후 예측에서 더 나은 결과를 내도록 합니다.
수학적 비유:
이를 더 수학적으로 설명하면, 모델은 각 샘플에 대해 확률 분포를 계산합니다. 여기서 좋은 샘플은 높은 확률을 부여받고, 나쁜 샘플은 낮은 확률을 부여받습니다. 그런 다음, 이 확률 분포를 기반으로 모델이 학습을 진행하면서 점점 더 높은 확률을 가진 샘플이 많이 선택되도록 조정됩니다.
이 과정은 마치 설문조사에서 선호도가 높은 선택지를 계속 선택하게끔 유도하는 것과 같습니다. 처음에는 다양한 선택지를 제공하지만, 시간이 지남에 따라 사람들이 더 많이 선택한 답변을 기준으로 더욱 정교한 선택을 하게 되는 방식입니다.
17. 피드백 루프(Feedback Loop):
ESM3는 단백질 구조의 예측과 생성을 반복적으로 학습하는 과정에서 피드백 루프를 형성합니다. 이 피드백 루프에서는 모델이 이전에 예측한 단백질을 평가하고, 그 결과에 따라 더 나은 예측을 하도록 피드백을 제공합니다. 이 피드백을 통해 모델은 점차 더 나은 성능을 얻게 됩니다.
비유:
이 과정을 요리사가 음식을 여러 번 만들면서 맛을 조금씩 조정하는 것에 비유할 수 있습니다. 처음에는 맛이 완벽하지 않을 수 있지만, 여러 번의 시도와 피드백을 통해 점점 더 맛있는 요리를 완성하게 됩니다.
18. 정렬을 통해 향상된 모델:
결과적으로, ESM3는 정렬 과정을 통해 처음보다 훨씬 더 정확하고 생물학적으로 유용한 단백질을 생성할 수 있습니다. 특히, 특정 생물학적 기능을 목표로 할 때, 이 모델은 기존의 단백질 데이터를 기반으로 새로운 단백질을 설계할 수 있으며, 이 과정에서 자연에 존재하지 않는 새로운 기능성 단백질을 개발하는 데 기여할 수 있습니다.
비유:
이 과정을 마치 스포츠 훈련에 비유할 수 있습니다. 처음에는 단순한 기술을 배우지만, 훈련을 통해 점차 더 복잡한 기술을 익히게 됩니다. 이처럼 ESM3도 초기에는 단순한 패턴을 학습하지만, 정렬 과정을 통해 더 복잡하고 정교한 단백질 구조와 기능을 예측하게 됩니다.
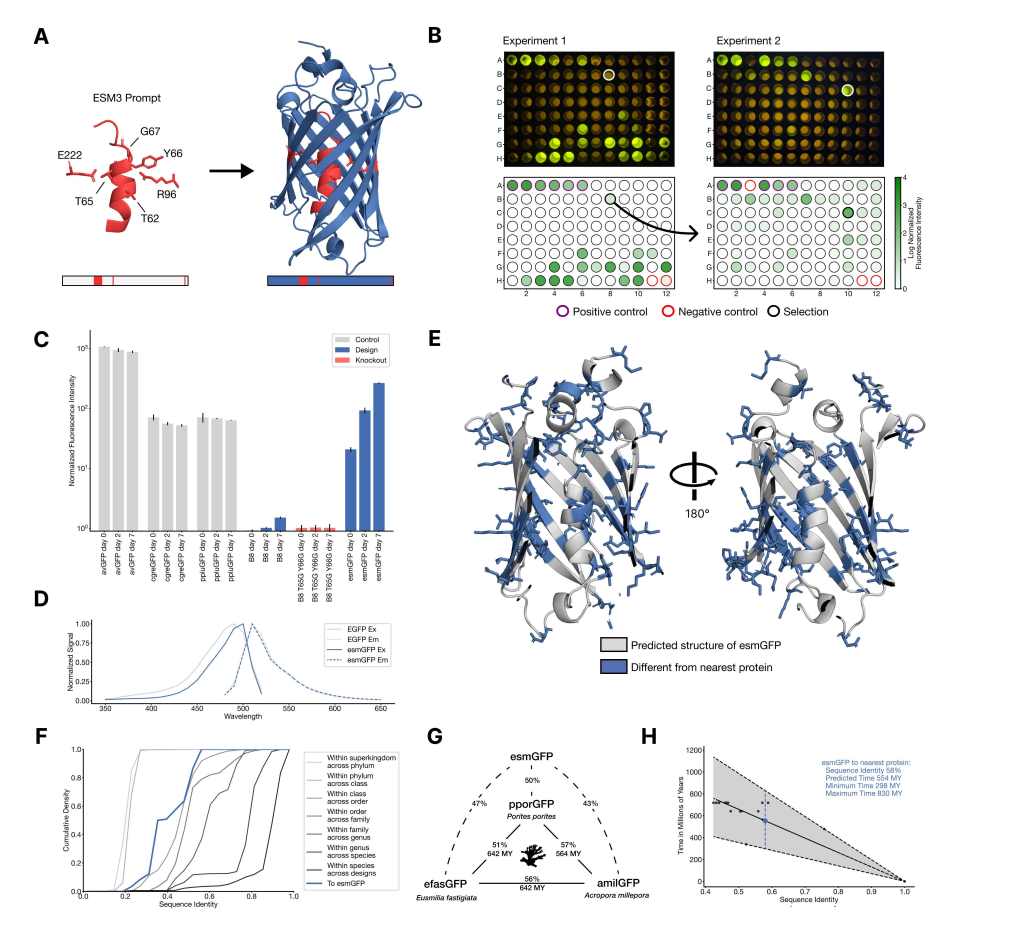
19. 새로운 형광 단백질의 생성 (Generating a New Fluorescent Protein):
이 논문에서 연구팀은 ESM3를 사용하여 전혀 새로운 형광 단백질을 생성하는 실험을 수행했습니다. 형광 단백질은 해파리나 산호가 빛을 내는 원리로 작동하는데, 생명과학 분야에서 특정 세포나 단백질을 가시화하는 데 중요한 도구로 사용됩니다.
형광 단백질의 생성 메커니즘:
ESM3는 주어진 조건 하에서 새로운 단백질 서열을 생성할 수 있습니다. 연구팀은 형광을 발현할 수 있는 중요한 아미노산 서열(예: 형광 발현에 중요한 특정 아미노산들)을 입력으로 주었고, ESM3가 이 서열을 바탕으로 전혀 새로운 형광 단백질을 생성하도록 했습니다.
비유:
이를 더 쉽게 설명하자면, 마치 특정 재료가 주어졌을 때 요리사가 그 재료를 바탕으로 새로운 요리를 창작하는 것과 같습니다. 연구팀은 형광 단백질의 기본 '재료'를 주었고, ESM3는 그 재료를 바탕으로 새로운 형광 단백질을 '요리'해냈습니다.
20. 형광 단백질을 생성하는 과정:
ESM3가 형광 단백질을 생성하는 과정은 단계적으로 이루어졌습니다.
1. 중요한 아미노산 서열 제공: 먼저, 형광 단백질에서 형광을 발현하는 데 중요한 아미노산 서열(예: Thr65, Tyr66 등)을 모델에 제공했습니다. 이는 마치 요리 레시피의 핵심 재료를 제공하는 것과 같습니다.
2. 구조적 중요 부위 지정: 형광 단백질의 일부 3차원 구조(중요한 알파 나선 구조 등)를 모델에 제공하여 ESM3가 단백질 구조를 예측하는 데 도움을 주었습니다.
3. 서열 및 구조 최적화: 그 후, ESM3는 이 입력 정보를 바탕으로 단백질의 전체 서열을 생성하고, 여러 번의 최적화 과정을 거쳐 최종 단백질 서열을 도출했습니다.
비유:
이 과정은 요리사가 새로운 요리를 만들기 위해 재료를 다듬고, 이를 반복적으로 시도하면서 점차 완성도를 높이는 과정과 유사합니다. 처음에는 간단한 재료와 구조만으로 시작하지만, 최종적으로는 완벽하게 조화로운 요리를 완성하게 되는 것입니다.
21. 새로운 단백질의 검증:
생성된 새로운 형광 단백질은 실험을 통해 검증되었습니다. 연구팀은 생성된 단백질을 대장균(E. coli)에 삽입하여 형광 발현을 테스트했습니다. 실험 결과, ESM3가 생성한 단백질 중 하나가 실제로 형광을 발현했으며, 이는 기존의 자연적인 형광 단백질과는 서열적으로 매우 멀리 떨어져 있었음에도 불구하고 성공적인 결과를 보였습니다.
비유:
이 과정은 새로운 레시피로 요리를 만든 후, 그 요리가 실제로 맛있고 기능하는지 확인하는 실험과 유사합니다. 모델이 생성한 단백질이 실제로 형광을 발현할 수 있었는지를 실험으로 확인하는 과정입니다.
22. 진화적 거리(Evolutionary Distance):
ESM3가 생성한 형광 단백질은 기존에 자연에서 발견된 형광 단백질과 서열적으로 58% 일치하는 것으로 나타났습니다. 이는 ESM3가 생성한 단백질이 기존의 자연 단백질로부터 매우 먼 진화적 거리에 위치한 단백질이라는 의미입니다. 연구팀은 이를 바탕으로, 이 단백질이 자연에서 발견되기까지 약 5억 년 이상의 진화가 필요했을 것이라고 추정했습니다.
비유:
이 과정을 이해하기 쉽게 설명하자면, 마치 어떤 고대 언어에서 새롭게 진화한 현대 언어와의 유사성을 찾는 것과 같습니다. ESM3가 생성한 단백질은 고대 자연에서 수백만 년에 걸쳐 진화한 단백질과는 다른, 새로운 '언어'를 사용하고 있는 것입니다.
23. 실험 결과와 결론:
결과적으로, ESM3는 자연에서 5억 년 이상 진화한 것과 동일한 수준의 서열 차이를 가진 새로운 형광 단백질을 생성할 수 있음을 보여주었습니다. 이는 단순히 기존의 데이터를 기반으로 예측하는 것을 넘어, 완전히 새로운 단백질을 생성할 수 있는 가능성을 입증하는 중요한 결과입니다.
계속해서 결론 (Discussion) 부분을 설명하겠습니다. 이 섹션에서는 ESM3 모델의 가능성과 그 의미에 대해 다루고 있습니다.
24. 결론 (Discussion):
연구팀은 ESM3 모델이 단백질 설계 분야에서 중요한 진전을 이루었음을 강조합니다. 특히, 자연의 진화 과정을 모방하여 새로운 단백질을 생성하는 능력은 생명과학 및 생물학 연구에서 매우 유망한 도구가 될 수 있습니다. 이 모델은 수억 년의 진화적 과정을 시뮬레이션하고, 그 과정을 통해 새로운 단백질을 효율적으로 설계할 수 있음을 보여주었습니다.
비유:
이 모델을 '자연의 설계자'라고 생각할 수 있습니다. 자연이 오랜 시간 동안 무작위 변이와 선택 압력을 통해 단백질을 최적화해왔듯이, ESM3도 비슷한 과정을 시뮬레이션하여 단백질을 생성합니다. 그러나 이 모델은 자연보다 훨씬 더 빠르게 진화의 가능성을 탐구하고, 기존의 자연적 경로와는 다른 새로운 단백질을 설계할 수 있습니다.
25. 언어 모델로서의 ESM3:
연구팀은 ESM3가 단백질 서열을 이해하고 생성하는 데 있어, 일반적인 언어 모델처럼 작동한다고 설명합니다. 단백질 서열은 마치 단어의 나열처럼 볼 수 있으며, 이 서열들이 상호작용하여 단백질의 구조와 기능을 결정합니다. 언어 모델이 문장을 이해하고 문법적으로 올바른 문장을 생성하듯이, ESM3는 단백질 서열을 학습하고 새로운 서열을 생성할 수 있습니다.
비유:
이를 문장 생성에 비유할 수 있습니다. 언어 모델이 단어들의 규칙과 패턴을 학습하여 문장을 생성하듯이, ESM3는 단백질의 서열, 구조, 기능 간의 규칙과 패턴을 학습하여 새로운 단백질 서열을 만듭니다. 그 결과, 새로운 단백질은 마치 자연에서 진화한 단백질처럼 생물학적 기능을 가질 수 있습니다.
26. 단백질 공간에서의 진화 (Evolution in Protein Space):
단백질의 변이와 진화는 특정 공간 안에서 이루어집니다. 이 공간에서는 각 단백질이 하나의 좌표로 나타내어지며, 이 좌표는 단백질의 서열, 구조, 기능을 나타냅니다. ESM3는 이 단백질 공간을 탐색하면서 기존에 존재하지 않았던 새로운 단백질을 생성할 수 있습니다. 이는 단백질이 자연에서 진화하는 경로를 시뮬레이션하는 것과 비슷한 방식으로 이루어집니다.
비유:
이를 '단백질 지도'에 비유할 수 있습니다. 이 지도에서 각 단백질은 하나의 위치에 있으며, 자연은 특정 경로를 따라 단백질을 진화시킵니다. ESM3는 이 지도를 탐색하며 자연에서 발견되지 않은 새로운 단백질 경로를 찾아낼 수 있습니다. 즉, 자연이 발견하지 못한 새로운 '길'을 만들어내는 것입니다.
27. 미래의 가능성 (Future Potential):
ESM3는 단백질 설계에서 더 많은 가능성을 열어줍니다. 이 모델은 현재 자연에서 발견되지 않은 새로운 단백질을 생성할 수 있는 능력을 갖추고 있기 때문에, 의약품 개발, 생명공학, 그리고 다른 생물학적 응용 분야에서 매우 유용할 수 있습니다. 또한, 단백질의 구조와 기능을 조합하여 특정 목적을 위해 최적화된 단백질을 설계하는 데 사용될 수 있습니다.
비유:
이를 마치 새로운 도구를 발명하는 과정에 비유할 수 있습니다. 기존의 도구들은 특정한 용도로 개발되었지만, 새로운 기술을 통해 더 효율적이거나 전혀 다른 용도의 도구를 발명할 수 있습니다. ESM3는 이러한 과정을 단백질 설계에 적용하여, 현재 존재하지 않는 새로운 생물학적 도구를 만들어낼 수 있습니다.
28. ESM3의 한계와 개선점:
연구팀은 ESM3 모델이 아직 초기 단계에 있으며, 향후 개선될 여지가 있음을 언급합니다. 예를 들어, 모델이 더 다양한 단백질을 생성할 수 있도록 데이터의 다양성을 늘리거나, 특정 생물학적 기능을 더 정확하게 예측할 수 있도록 정교한 조정이 필요할 수 있습니다. 또한, 실험적 검증을 통해 모델이 생성한 단백질의 실제 기능을 확인하는 과정이 더 많이 필요합니다.
비유:
이를 마치 새로운 기술을 개발한 후, 그 기술을 상용화하기 위해 여러 단계의 테스트와 개선이 필요한 과정에 비유할 수 있습니다. 새로운 기술은 처음에는 한계가 있을 수 있지만, 지속적인 연구와 개선을 통해 더욱 완벽해질 수 있습니다. ESM3도 마찬가지로, 향후 더 많은 데이터와 실험을 통해 완성도를 높여갈 수 있습니다.
