✅ 항체의 구조 및 항체 엔지니어링의 기본 개념
🖐 Immunoglobulin (Ig, 항체, 면역글로불린)
면역계가 침입자를 인식하고 제거하기 위해 생성하는 단백질
🖐 Light Chain (LC)
가벼운 사슬
항체의 한쪽 팔을 구성
🖐 Heavy Chain (HC)
무거운 사슬
항체의 중심 축과 팔을 형성
항체는 Y자 모양의 단백질로, 두 개의 같은 Light Chain과 두 개의 같은 Heavy Chain이 붙어 만들어짐
이 네 개의 체인이 모여 하나의 완전한 항체를 구성
🖐 Heterodimer
서로 다른 두 단백질(여기선 HC + LC)이 짝을 이룸
🖐 Disulfide bond (이황화 결합)
단백질 사이를 안정하게 연결해 주는 화학 결합
한 개의 LC + HC가 짝을 이루고,
같은 구조가 하나 더 있어 총 2쌍 = 4개의 체인이 결합해 완전한 항체(Y자 구조)가 됨
이 결합은 화학적으로 이황화 결합으로 묶임
🖐 Domain
단백질 내에서 기능적으로 독립적인 구조 단위
🖐 CL (Constant Light domain)
변하지 않는 영역
구조적 안정성 제공
🖐 VL (Variable Light domain)
변이 영역
항원 인식에 관여
🖐 Isotype
항체의 종류 (IgA, IgD, IgE, IgG, IgM)
기능과 위치가 다름
Light Chain은 CL + VL, 두 개의 도메인으로 구성
Heavy Chain은 isotype 마다 도메인 수가 다르며, 항체의 종류(역할과 위치)를 결정
🖐 항체 종류와 기능
| 항체 종류 | 도메인 수 | 기능/특징 |
|---|---|---|
| IgG, IgA, IgD | V + 3C (총 4개 도메인) | 일반적인 면역 반응, IgG는 혈중 가장 많음 |
| IgM, IgE | V + 4C (총 5개 도메인) | IgM은 초기 면역 반응, IgE는 알레르기 반응에 관여 |
| IgA, IgM | J-chain 있음 | 다량체 형성 -> IgA(2개), IgM(5개)로 연결되어 작동 |
| IgG, IgD, IgE | Monomer 형태 | 각각 독립적인 항체로 작동 |
🖐 IgG1
가장 대표적인 항체 종류
치료용 항체로 가장 많이 사용됨
🖐 Framework
항체의 기본 골격 구조
✅ 항체 엔지니어링
| 전략 | 설명 |
|---|---|
| Effector function engineering | Fc 영역을 조절하여 면역세포 활성화 정도를 조절 |
| Antibody humanization | 마우스 항체를 인간 항체로 바꿔 면역원성 낮춤 |
| Affinity modulation | 항원 결합 친화도를 높이거나 조절 |
| Stability enhancement | 구조적 안정성 및 저장성 향상 |
🖐 항체의 전체 구조
| 구조 | 설명 | 그림에서 색 |
|---|---|---|
| Fab (Fragment antigen-binding) | 항원에 결합하는 부위 각 팔 끝에 하나씩 존재 | 왼쪽과 오른쪽 팔 부분 |
| Fv (Variable region) | Fab의 일부 항체마다 다른 항원 특이성을 결정 (VH + VL) | |
| Fc (Fragment crystallizable) | 면역세포와 결합하여 신호를 전달해, 면역 반응 유도 | 하단 중심 부분 |
| Hinge | Fab와 Fc를 연결하는 유연한 부분 | 가운데 접힌 부위 (노란색 이황화 결합) |
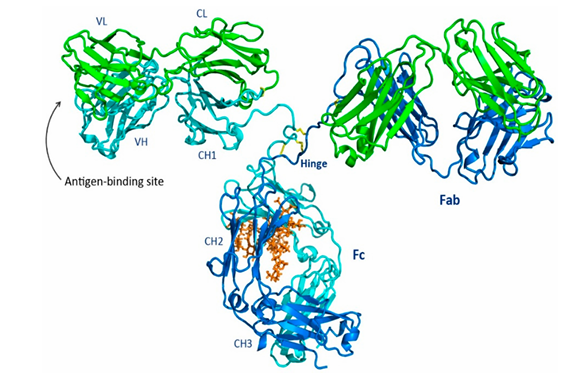
- Green = Light Chain (LC)
- Cyan/Blue = Heavy Chain (HC)
- Orange sticks = Glycan (당) → Fc에 붙어 면역 기능에 영향
- Yellow sticks = Interchain disulfides (사슬 간 이황화 결합)
🖐 도메인 구조 및 접힘 양상
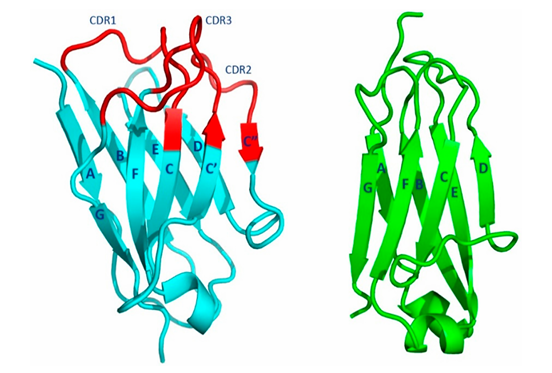
🔷 왼쪽 그림: VH 도메인 (cyan + red)
VH = Heavy chain의 Variable domain
CDR1, CDR2, CDR3 (빨간색): 항원과 결합하는 부위 (Complementarity Determining Regions)
나머지 부분(cyan)은 안정적인 구조 형성 → scaffold 역할
🟩 오른쪽 그림: CL 도메인 (green)
CL = Light chain의 Constant domain
C 도메인은 구조적으로 안정적이며 항체 골격 유지에 중요
✅ 도메인 구성 및 접힘 방식
항체는 여러 domain 으로 구성
이 도메인 하나하나가 비슷한 방식으로 접혀서 안정적인 구조를 만들어냄
이 접힘 방식을 "면역글로불린 접힘 (Immunoglobulin fold)"이라고 부름
🖐 Immunoglobulin fold 란?
모든 도메인(VH, VL, CH, CL)은 공통적으로 약 110개의 아미노산으로 구성되며, 고유한 접힘 구조를 가짐
→ 이를 "면역글로불린 접힘 (Immunoglobulin fold)"이라고 함.
이 도메인은 두 개의 얇은 판처럼 생긴 β-sheet (베타 시트) 두 장으로 이루어져 있고,
이 두 장이 서로 마주 보면서 샌드위치처럼 붙어 있음
각각의 β-sheet는 여러 가닥(β-strand)으로 구성
각 β-strand는 화살표 모양으로 그려지고, 그 방향은 아미노산 사슬이 접히는 방향을 나타냄
✅ C 도메인 (CL, CH1~CH3)의 구조
✅ β-sheet 구조
첫 번째 β-sheet: A, B, E, D (총 4개 가닥)
두 번째 β-sheet: C, F, G (총 3개 가닥)
이 두 시트가 딱 붙어 있으면서 안정적인 베타 샌드위치 구조를 이룸
✅ 이황화 결합
두 시트를 Cysteine (시스테인)이라는 아미노산이 공유결합인 S-S 결합(disulfide bond)으로 연결
이 결합은 접힘을 안정화
→ 도메인 내부를 고정시켜주는 핵심 결합
ex) VH, CH1, CL 등 각 도메인의 안에서 B-strand와 F-strand 사이에 있음
✅ 비공유결합 (Non-covalent interactions)
두 β-sheet가 서로 마주 보는 면에 있는 side chains 사이에 다음과 같은 비공유적 상호작용이 일어남:
-
수소결합 (Hydrogen bonding)
-
소수성 상호작용 (Hydrophobic interactions)
-
반데르발스 힘 (Van der Waals)
-
이온결합 (Electrostatic interactions)
→ 이런 결합들이 β-sheet 샌드위치를 단단히 붙잡아줌
✅ V 도메인 (VL, VH)의 구조
C 도메인보다 조금 복잡함
항체의 V 도메인은 항원과 직접 결합해야 하니까 더 다양하고 유연해야 함
항체의 V 도메인(VL, VH)은 light chain(LC)과 heavy chain(HC)의 N말단 (N-terminus)에 있음
이 V 도메인이 바로 항원 결합 부위 (antigen binding site)를 형성
✅ β-sheet 구조
첫 번째 시트: A, B, E, D (C 도메인과 동일)
두 번째 시트: C'', C', C, F, G
C’과 C’’이라는 가닥이 C와 D 사이에 삽입되어 더 많음
구조가 더 복잡해지고, 결과적으로 더 긴 loop들이 생김
그래서 더 유연하고, CDR 루프를 만들 수 있는 구조적 여유가 생김
이 loop들이 바로 CDR (항원 결합 루프)의 위치가 됨
V 도메인은 보통 덜 조밀하며, 유연성이 큼
| 항목 | 설명 |
|---|---|
| 구조 형태 | 2개의 anti-parallel β-sheet로 구성된 β-sandwich |
| 결합 방식 | B-strand와 F-strand 사이 이황화 결합 (disulfide bridge) |
| C 도메인 구조 (예: CL) | • β-sheet 1: ↓A ↑B ↓E ↑D (4-strand) • β-sheet 2: ↓C ↑F ↓G (3-strand) |
| V 도메인 구조 (예: VH, VL) | • β-sheet 1: ↓A ↑B ↓E ↑D • β-sheet 2: ↓C’’ ↑C’ ↓C ↑F ↓G (5-strand) • CDRs는 β-strand 사이의 루프에 위치 |
C vs V 도메인의 차이
| 항목 | C 도메인 | V 도메인 |
|---|---|---|
| 루프 길이 | 짧음 (안정성 중심) | 길고 유연 (항원 인식 중심) |
| β-strand 수 | 4 + 3 (총 7) | 4 + 5 (총 9) |
| 기능 | 구조적 골격 제공 | 항원 결합 부위 제공 |
✅ 항체의 Fab 영역 구조, 항원결합 부위의 형성 방식과 elbow angle
🖐 Fab(Fragment antigen-binding) 영역이란?
항체의 한쪽 팔에 해당하며, 항원을 인식하고 결합하는 핵심 영역
하나의 Fab은 다음 도메인들로 구성됨:
-
Light Chain (LC): VL + CL
-
Heavy Chain (HC): VH + CH1
이 두 영역이 합쳐져 하나의 Fab 형성
✅ 항원결합부 형성
VL과 VH가 짝을 이루어 항원결합부 (antigen-binding site, Fv region)를 형성
VL–VH 쌍은 각각 β-sheet 구조를 이루고, 다음처럼 구성
항원결합 도메인의 β-sheet 구조: ↓C″ ↑C′ ↓C ↑F ↓B
→ 이 β-strand들이 통처럼 접히며, 사이의 loop가 CDR 로 노출됨
반면, CL–CH1 도메인은 서로 다른 방향에서 접근하여
↓A ↑B ↓E ↑D β-sheet 구조를 기반으로 직각(수직)에 가깝게 단단히 붙음
CL과 CH1은 서로 마주 보는 β-sheet 면을 가짐
↓A ↑B ↓E ↑D는 β-sheet의 네 개 strand를 뜻함
이 β-strand들은 anti-parallel 구조로 한 면의 평평한 시트(face)를 형성
CL과 CH1 각각의 도메인은 이런 시트를 가지고 있고, 이 시트끼리 마주 보면서 붙음
🖐 Elbow Bend(= Elbow Angle) 란?
Fab 안의 VL–VH 도메인 쌍(Fv 영역)과 CL–CH1 도메인 쌍 사이의 각도를 뜻함
VL–VH, CL–CH1 각각은 쌍으로 거의 대칭 구조를 이룸
이 두 쌍이 이루는 가상의 축들 사이의 각도 = elbow angle(elbow bend)
Fab 전체 구조의 접힘 모양에 영향
항체가 항원에 접근할 수 있는 공간성 조절
🖐 Switch region 이란?
“V 도메인과 C 도메인을 연결하는 구조적인 연결 부위”를 의미
몇 개의 아미노산으로 구성된 유연한 폴리펩타이드 서열
항체의 Fab 영역을 구성하는 VH–CH1, VL–CL 사이의 경계 지점
도메인 사이를 물리적으로 연결
일반적으로 항체 접힘 구조에서 유연성이 가장 큰 부분 중 하나
-> Fab 도메인이 약간의 움직임을 할 수 있게 해줌
| 용어 | 위치 | 의미 | 기능 |
|---|---|---|---|
| Switch region | V 도메인과 C 도메인 사이의 연결부위 (VH–CH1 또는 VL–CL 사이) | 실제 연결을 해주는 유연한 아미노산 사슬 | 도메인 간 유연성 제공 |
| Elbow bend (angle) | V 도메인 쌍(VH–VL)과 C 도메인 쌍(CH1–CL) 사이의 기하학적 각도 | 두 도메인쌍 간의 상대적인 기울기 | 항체 구조의 입체적 형태 결정 |
VL──VH ← Fv 도메인 쌍 (항원 인식 부위)
║
switch region ← 유연한 연결 부위 (Switch)
║
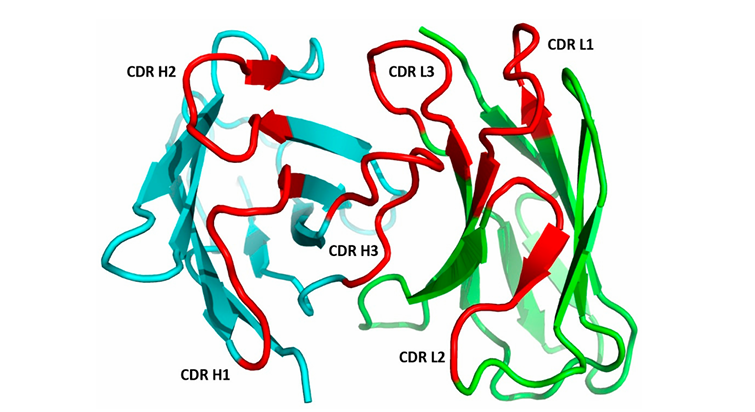
CL──CH1 ← Constant 도메인 쌍✅ Fv 영역과 CDR 구조
🖐 Fv (Fragment variable)
항체의 VH(Heavy chain variable) + VL(Light chain variable) 도메인이 결합된 부분
이 부분이 N 말단 region 과 결합하면서 항원 결합 부위 구성
여기에 항체-항원 결합의 핵심 부위인 CDR 6개가 모여 있음
CDR-L1, L2, L3 (light chain에서 3개)
CDR-H1, H2, H3 (heavy chain에서 3개)
이 6개 루프가 서로 가까이 배치되어 항체의 결합부위 (paratope)를 형성.
🖐 N-terminal (N 말단)
단백질 서열의(항체) 시작 지점
단백질은 아미노산이 일렬로 연결된 폴리펩타이드이고,
항상 한쪽 끝은 아미노기(-NH₂), 다른 쪽 끝은 카복실기(-COOH)로 끝남
| 끝 | 이름 | 설명 |
|---|---|---|
| 시작점 | N-terminal (N 말단) | 아미노산 사슬이 시작하는 쪽. 첫 번째 아미노산이 있는 자리. |
| 끝점 | C-terminal (C 말단) | 마지막 아미노산이 있는 쪽. 사슬이 끝나는 자리. |
[ N-term ] — VL — CL — [hinge] — CH2 — CH3 — [ C-term ]
VH와 VL 도메인 (즉, Fv region)은 항체의 N-terminal 쪽에 위치
✅ CDR 위치 및 구조
| CDR 이름 | 위치한 β-strands |
|---|---|
| CDR-L1, H1 | ↑B–↓C 사이 |
| CDR-L2, H2 | ↑C′–↓C″ 사이 |
| CDR-L3, H3 | ↑F–↓G 사이 |
이 구조는 항체 입체 구조의 기준틀인 β-sheet 위에 loop 형태로 돌출됨
이 루프들이 바로 CDR이고, 이곳이 항원과 접촉해 인식함
🖐 SDR (Specificity-Determining Residues)
SDR은 항체가 항원을 인식하는 데 실제로 관여하는 CDR 내부 잔기들
CDR 전체가 아닌, 그중에서도 항원과 직접 상호작용하는 핵심 잔기들을 말함
✅ 항원 종류에 따른 항체 결합 방식
항체가 인식하는 항원 종류에 따라 결합 부위 모양이 달라짐
| 항원 종류 | 결합 부위 모양 | 특징 |
|---|---|---|
| Hapten (소분자) | 깊고 좁은 pocket | 항원 결합 부위가 깊고 폐쇄적 |
| Peptide (펩타이드) | 홈처럼 생긴 groove | 길쭉한 홈 모양, 펩타이드가 그 안에 끼워짐 |
| Protein (단백질) | 넓고 평평한 면 (surface) | 접촉면이 넓고 복잡함 |
이런 구조적 특징을 바탕으로, 항체 라이브러리를 항원 종류별로 최적화 가능
✅ 항체 다양성의 분자적 기원
VH 도메인: V, D, J 유전자 조합
VL 도메인: V, J 조합
여기에 체세포 고돌연변이(somatic hypermutation)까지 더해져, 항체의 CDR 서열 다양성이 무한에 가까움
✅ 항체–항원 결합 시 구조적 변화
✅ 1. Lock-and-Key (자물쇠-열쇠 모델)
| 특징 | 설명 |
|---|---|
| 구조 변화 | 거의 없음 |
| 의미 | 항체와 항원이 이미 딱 맞는 모양을 가지고 있어서 결합할 때 구조 변화가 거의 없음 |
| 예시 | 열쇠가 자물쇠에 딱 맞듯이, 두 단백질이 기존 형태 그대로 결합 |
unbound (결합 전)와 bound (결합 후) 상태의 구조가 거의 동일
주로 backbone (주사슬) 구조가 그대로 유지됨
✅ 2. Induced Fit (유도 적합 모델)
| 특징 | 설명 |
|---|---|
| 구조 변화 | 매우 큼 |
| 의미 | 결합하면서 서로의 모양이 변화됨 –> 마치 손과 장갑처럼 딱 맞게 조정되는 느낌 |
| 변하는 부위 | 특히 CDR (complementarity determining region) 중 CDR-H3가 가장 자주 변함 |
항체나 항원의 side chain 뿐만 아니라 backbone 까지 변형됨
VL-VH 간의 상대적인 방향도 달라질 수 있음
심지어 Fab elbow angle도 변할 수 있음
이 메커니즘 덕분에 항체는 더 다양한 항원에 적응 가능 (plasticity)
✅ 3. Conformational Selection (구조 선택 모델)
| 특징 | 설명 |
|---|---|
| 구조 변화 | 결합 전 이미 다양한 구조 존재 |
| 의미 | 항원 또는 항체가 다양한 구조 중 일부를 미리 갖고 있고, 그중에 항체가 선택적으로 결합 |
| 포인트 | 결합이 항체의 유도보다는 항원이 미리 준비한 구조 중 하나를 선택하는 방식 |
항원은 다양한 상태(구조)를 자연적으로 샘플링하고 있음
항체는 그 중 결합에 유리한 구조를 선택해서 결합
항원의 구조는 주변 환경 (microenvironment)에 따라 달라질 수도 있음
🖐 CDR의 Canonical Structures란?
루프의 길이, 형태, 특정 보존 아미노산에 의해 결정되는 대표적인 입체 구조 유형
CDR 루프 구조가 예외 없이 다양할 거라는 기존 믿음과는 달리,
실제로는 대부분이 몇 가지 정형화된 구조(= Canonical Structures)만 가짐
즉, 항체마다 구조가 완전히 독특하지는 않음
✅ Canonical Structure의 결정 요소
- 루프의 길이
- 접힘 패턴(conformation, backbone 방향성)
- 보존된 아미노산 패턴(특정 위치에 존재하는 Gly, Pro 등의 잔기 + 주변 FR 영역까지 포함)
특히, CDR의 길이가 항원 결합 부위(topography, 항체 결합 부위의 표면 지형)의 주요 결정 요소라는 것이 밝혀짐
루프의 길이가 곧 canonical structure(대표적인 구조 유형)를 결정하기 때문
✅ 1. CDR 루프 길이 → 구조 결정
루프는 길이에 따라 접힘 패턴(conformation)이 다름
특히 길이가 짧은 루프는 구조가 제한되어 있지만,
길어질수록 유연성과 가능한 구조가 증가함
ex)
CDR-L1이 11개의 아미노산이면 특정 구조군에 속할 확률이 매우 높음
반면 14개의 아미노산이면 다른 구조군에 속할 수 있음
따라서 루프의 길이는 곧 그 루프의 canonical structure를 결정하게 되는 주요한 기준
✅ 2. Canonical structure → 항원 결합부 topography 결정
루프들이 어떤 방향으로 휘고, 얼마나 튀어나오는지에 따라
항원이 인식하는 항체의 표면 구조가 달라짐
-> 이게 바로 topography (표면지형)
ex)
짧은 루프들만 있는 항체: 평평하고 좁은 결합부
길고 유연한 루프: 홈(groove) 구조나 깊은 포켓이 생김
CDR 루프의 길이 → 루프 구조 (canonical structure) → 결합부 지형(topography)
이 요소들을 통해 항체 구조를 예측하거나 모델링할 수 있게 됨.
✅ CDR 별 구조 정형화 여부
| CDR 구역 | Canonical 구조 존재 여부 |
|---|---|
| CDR-L1, L2, L3 | 구조 정형화됨 |
| CDR-H1, H2 | 구조 정형화됨 |
| CDR-H3 | 다양한 길이와 서열 가짐, 예측 어려움 |
H3는 길이 다양성도 크고, 항원 특이성에 핵심이라서 가장 유연함
-> 다른 CDR 들은 어느정도 정형화 되어 있으나, HCDR3는 정형화 되어 있긴 하나 다른 CDR들 보다는 매우 다양함
✅ 최신 연구 동향
더 많은 고해상도 항체 구조가 확보되면서,
알고리즘적으로 CDR 구조 클러스터링을 수행하고 있음
CDR 관련 정보 단백질 데이터 뱅크(PDB)의 항체 구조
https://dunbrack2.fccc.edu/pyigclassify2/
✅ CDR-H3의 구조적 특성과 중요성
CDR-H3 루프를 두 구간으로 나누어 설명
| 구간 | 설명 |
|---|---|
| Torso | 루프가 시작하는 부분 (framework 근처) |
| Head | 루프가 위로 솟는 상단 부분 (항원과 직접 접촉 부위) |
각 구간은 서로 다른 방식으로 정형화된 구조를 가질 수 있음
✅ Fc Region
항체(IgG)를 단백질 분해효소(papain) 로 절단하면 세 개의 조각으로 나뉘는 것이 관찰됨
이 중 하나의 조각은 여전히 항원을 인식하고 결합할 수 있었음
→ 이것이 Fab(fragment antigen-binding) 이라고 부르는 조각
Fc는 항체의 하단부에 위치한 영역으로, 두 개의 CH2 및 CH3 도메인으로 구성됨 (각각의 heavy chain에서 나옴)
항체의 효력 기능(effector function) 대부분이 Fc에서 수행됨
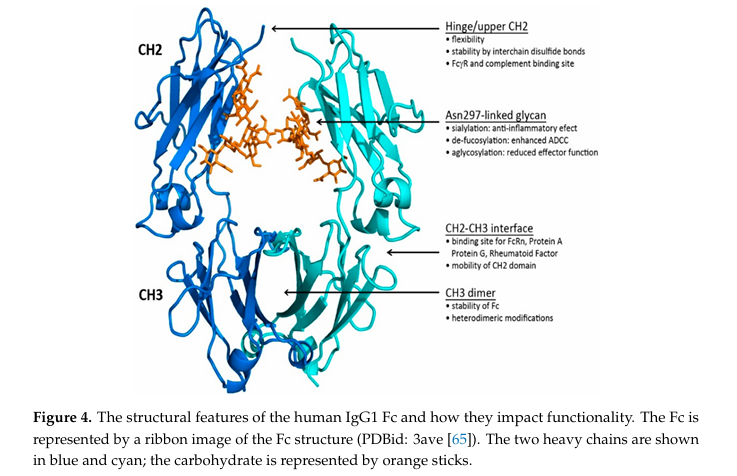
파란색과 청록색: 두 개의 heavy chain (CH2, CH3)
주황색 스틱: 당쇄 (glycan), Asn297에 연결됨
✅ 구조적 구성 요소 및 기능
1️⃣ CH2 도메인 (상단부)
서로 직접 접촉하지 않음 → 유연성 가짐
Asn297 에 부착된 당(glycan) 구조가 CH2 사이 공간을 채움
이 당은 수소 결합을 통해 서로 상호작용할 수 있고, CH2의 유연성(flexibility) 을 유도함.
CH2에 부여된 유연성은 C1q 및 FcγR과의 상호작용에 중요
2️⃣ CH3 도메인 (하단부)
서로 단단히 붙음 → Fc의 안정성에 기여
CH3 dimer 형성: 항체 구조 유지에 중요
✅ 주요 기능
Fc는 Fc 수용체(FcγR) 및 보체 단백질 C1q와 상호작용하여
- ADCC (항체의존 세포독성)
- CDC (보체의존 세포독성)
- ADCP (항체의존 식세포작용)
- Trogocytosis (세포막 교환 현상)
- 매개물 분비 유도
- opsonized 입자의 엔도사이토시스
- FcRn과의 상호작용을 통해 항체의 반감기 조절 관여
Fc는 다양한 면역세포(monocyte, NK cell 등) 와 작용하므로,
Fc의 기능 조절을 위한 엔지니어링이 활발함
✅ Hinge region
✅ Hinge의 위치와 역할
Hinge는 CH1과 CH2 도메인 사이의 영역으로, 두 개의 Fab와 Fc 사이를 연결함.
기능적으로 Fab에 유연성을 부여, 여러 항원 종류에 결합 가능하게 함.
Fc는 면역 체계의 다른 구성 요소들과 독립적으로 상호작용할 수 있도록 해줌
✅ Hinge의 세 부분: Upper, Core, Lower
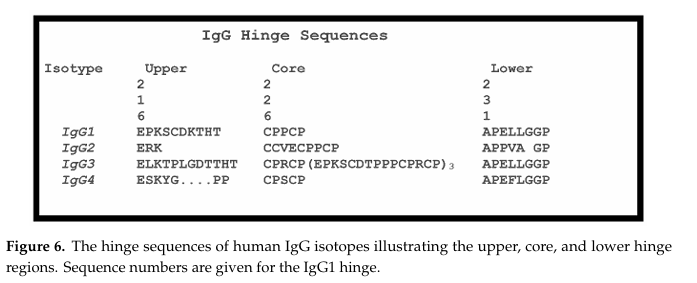
| 구분 | 기능 |
|---|---|
| Upper hinge | Fab의 회전과 움직임을 허용 (N 말단에 위치) |
| Core hinge | HC 간 이황화결합(Cys) 포함 → 안정화 |
| Lower hinge | Fab에 대한 Fc 움직임을 허용 (C 말단에 위치), FcγR 결합에 관여 |
이 3가지 영역은 IgG1~4 마다 길이와 Cys 수가 다름 → 항체의 구조 및 안정성 변화 유발.
특히 core hinge의 Cys 수는 Fc 안정성과 Fab-arm exchange에 영향.
🔎 ex)
IgG4: 유일하게 Fab-arm exchange가 자연적으로 일어남 → bispecific 항체 형성 가능.
✅ hinge engineering의 중요성
hinge는 protease(단백질 분해 효소) 공격에 민감함
특히, hinge 절단으로 인해 항체가 클리핑(clipping) 되면 Fc 효능(CDC, ADCC 등) 소실 위험 있음.
hinge 절단을 막기 위해 돌연변이(mutagenesis) 활용 → protease 저항성 부여 가능.
이런 변화는 염증 조직에서 치료 항체의 안정성을 높임.
이 외에도 hinge는 전체 항체의 구조적 유연성, 안정성, effector 기능 조절에 매우 중요.
✅ 구조 기반 항체 엔지니어링 전략
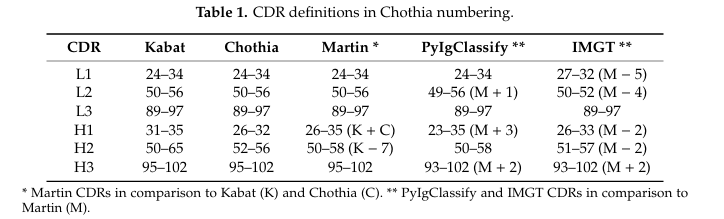
✅ Kabat 정의
최초의 CDR 정의 방식
다양한 항체 서열을 비교하여 변이성이 높은 구간을 CDR로 지정
👉 구조보다 서열 기반
✅ Chothia 정의
X-ray 결정 구조를 기반으로 함
항원 결합 루프의 공간적 구조에 따라 CDR 경계를 정의
👉 canonical structure 개념(보존된 루프 구조)을 도입
✅ Martin 정의
Kabat과 Chothia의 혼합형
실제로 항원과 접촉하는 잔기 중심으로 CDR 경계 설정
👉 실질적으로 결합에 관여하는 구조적 잔기만 포함하는 정의
ex) H2는 Kabat보다 7개 짧음 → 접촉 없는 루프 생략
✅ PyIgClassify
Martin 기준 기반
N-말단 방향에 L2 (+1), H1 (+3), H3 (+2) 잔기 추가
👉 대칭적이고 정밀한 구조 분류에 유리
✅ IMGT 정의
가장 체계적인 항체 전용 데이터베이스 (IMGT) 기반
TCR, MHC 등도 포함 가능한 범용적 정의
구조/서열 일관성 유지에 유리하지만 항원 결합 잔기를 일부 누락할 수 있음
✅ Humanization 관점에서의 해석
CDR 경계 설정 시 고려사항
- CDR은 가능한 짧게 설정해야 함 → 비인간 유래 잔기 최소화
- 항원과 접촉하는 모든 잔기는 포함되어야 함
👉 따라서, Martin 정의는 적절한 절충안 (binding 잔기 포함 + 비접촉 루프 생략)
Chothia numbering 은 구조 기반으로 insertions/deletions 보정이 잘 됨
👉 align & 비교에 유리
👉 대부분의 구조 기반 연구는 Chothia 기준 사용
🛑 But
실제 항원 결합 잔기의 20%는 CDR 외부 (FR 내)에 있음
어떤 경우에는 이 FR 잔기들이 binding에 더 중요
-> 이 FR 잔기의 backmutation 이 필요
✅ Germline 선택 기준
1️⃣ 전체 V영역 유사성 기준
VH 또는 VL 전체 서열을 기준으로 가장 유사한 human germline 선택
2️⃣ FR만 비교 (가장 많이 쓰임)
CDR은 어차피 graft되므로 → FR scaffold가 유사하면 구조 보존됨
3️⃣ CDR 유사성 기반 선택 (super-humanization)
canonical structure에 의존하여 CDR 구조 유지를 목표로 germline 선택
또한, 실험 결과 상
단순한 서열 유사성만 보고 human germline을 고르면 결합능 손실 가능성 있음
오히려 CDR의 구조적 호환성 (canonical structure)이 더 중요할 수 있음
✅ VH–VL interface residue 보존의 중요성과 Germline 선택 기준
항체 가변 영역의 아미노산들을 Chothia numbering 기반으로 표시
| 색상 | 의미 |
|---|---|
| 🔴 빨간색 원 | CDR (Complementarity-Determining Region) – 항원 결합 부위 |
| ⚪ 회색 원 | Vernier zone residues – CDR을 지지하는 framework 잔기 |
| 🟢 초록색 원 | VH–VL interface residues – VH와 VL 간 상호작용에 직접적으로 관여 |
IGHV1-69*01
▶ VH
Interface residues (7개): 초록색 원대부분 FR2에 위치, 일부는 CDR3 바로 근처
IGKV4-1*01
▶ VL
Interface residues (8개): 초록색 원역시 대부분 FR2, 일부는 CDR3 근처
CDR3를 둘러싼 2개의 잔기를 제외한 나머지 모든 잔기는 VH와 VL 모두의 FR2에 존재
✅ VH–VL interface 보존의 중요성
항체 humanization 시 가장 흔한 전략은 CDR을 human germline 프레임워크에 graft하는 것.
하지만 이때 VH–VL 간 orientation(방향)이 원본 항체와 다르게 바뀌면 antigen에 결합하지 못할 수 있음.
VH와 VL이 서로 어떤 각도로 결합해 있는지를 말함.
즉, 두 chain 간의 상대적 위치와 회전 방향
🔎 ex)
마우스 항체 HyHEL-10을 인간화했더니,
항원에 대한 결합력(affinity)이 10배나 감소함.항원과의 직접 상호작용(CDR)은 그대로 유지되었음.
그런데도 문제가 발생한 이유는,
→ VH와 VL이 결합하는 상대적 방향(orientation)이 바뀌었기 때문임.마우스 항체에서 VH의 39번(K)과 47번(Y)은 독특한 잔기였지만,
인간화하면서 보편적인 잔기 Q, W로 바뀜.이게 VH와 VL의 결합 방향을 바꿔버렸고,
→ 결과적으로 항원과 결합하는 CDR의 입체 배치가 바뀜.
이 두 개의 잔기를 다시 원래대로 복원(back-mutation)하자,
→ 결합력이 완전히 회복됨.
✅ VH–VL orientation 이란?
단순한 결합 유무가 아님
VH와 VL은 물리적으로 항상 붙어 있지만, 어떻게 붙어 있느냐가 중요
이걸 VH–VL orientation 또는 packing angle이라고 함
VH와 VL이 어떤 회전 각도, 기울기, 위치로 붙어 있느냐에 따라
그 위에 위치한 CDR (특히 CDR-H3)의 공간 구조가 달라짐
그래서 VH–VL 결합 orientation이 항원 결합력에 결정적인 영향을 줌
✅ Germline 선택 시 고려 요소
| 요소 | 설명 |
|---|---|
| (1) 안정한 VH–VL 형성 | 물리적으로 잘 맞물리는 VH–VL 쌍 선택 |
| (2) parental orientation 유지 | 기존 마우스 항체의 VH–VL 방향 유지를 우선 고려 |
| (3) interface residue 보존 | 특히 interface 상 잔기들은 human germline에서 동일한 아미노산을 가진 쌍을 선택하거나 back-mutation 고려 |
1️⃣ Germline pairing 통계
인간 항체 repertoires는 매우 다양한 VH–VL pairing을 생성함.
대부분 랜덤처럼 보이지만, 일부 VH–VL 조합은 선호되어 더 자주 사용됨.
단, packing angle 자체는 germline보다 CDR-H3 구조에 더 영향을 받음.
2️⃣ VH–VL pairing에서 무엇이 pairing을 결정하는가?
CDR-H3가 결정적
VH–VL 인터페이스 중 상당 부분이 CDR-H3에 의해 형성됨.
CDR-H3는 매우 다양하고 구조적으로 유연함 → VH–VL 결합 방식에 큰 영향을 줌.
그래서 같은 VH–VL 조합이라도, CDR-H3의 형태에 따라 결합의 안정성이나 방향이 달라질 수 있음.
즉, pairing의 성능은 CDR-H3 서열/구조에 따라 달라지고,
이 때문에 모든 germline 조합의 친화도 차이를 표로 정리하기 어려움
3️⃣ VH–VL 조합에 따라 안정성(Tm)이 다름
🔎 ex)
고정된 CDR-H3를 가진 상태에서,
4개의 heavy chain germline (VH) × 4개의 light chain germline (VL) 조합을 만들어 총 16개의 Fab을 제작각 조합에 대해 Tm (melting temperature)를 측정 → 항체 안정성 비교
같은 CDR-H3를 가지고 있어도 VH–VL germline 조합에 따라 최대 20도 이상 차이가 남
→ VH–VL pairing 자체가 항체의 안정성에 엄청난 영향을 줄 수 있음
4️⃣ Humanization 시 적용되는 시사점
“좋은 pairing을 고르면, humanization 후에도 안정성과 친화도가 유지될 수 있다”
humanization 과정에서 대부분 sequence similarity(즉, mouse 항체와 얼마나 비슷한가)만 고려
그러나 실제로는 VH–VL interface 상에 있는 몇 개의 잔기들이 구조적으로 훨씬 더 중요함
5️⃣ VH–VL interface residue 보존
같은 CDR을 graft하더라도,
VH–VL interface의 구조가 보존되지 않으면
항체의 구조가 무너지거나, orientation이 틀어지거나, 안정성이 떨어질 수 있음.
6️⃣ 그래서 어떻게 해야 하나?
CDR은 그대로 graft하되,
VH–VL interface 잔기만은 parental(mouse) 항체에서 그대로 유지하거나, 보존된 human germline을 선택
CDR은 그대로 이식되므로, 소수의 FR 위치만 고려하면 됨
✅ VH–VL 인터페이스 기반 humanization 전략
CDR-H3는 고정되어 있다고 가정하고,
FR2 기반 interface 잔기 7개 (VH) + 8개 (VL)을 확인함.
human germline에서 해당 잔기들이 동일하지 않다면
해당 germline은 배제하거나
back-mutation으로 원래 잔기를 복원함.
항체 humanization을 잘 하려면,
단순히 CDR을 human germline에 graft하는 것만으로는 부족하고,
VH–VL의 orientation (packaging 방향)까지 유지되어야 안정성과 항원 결합력을 보존할 수 있음
| 상황 | 예측 도구가 할 수 있는 일 |
|---|---|
| 여러 human germline 후보 중 선택해야 할 때 | VH–VL interface 구조적으로 가장 잘 맞는 pair 추천 |
| humanization 후 VH–VL packing이 바뀌었는지 알고 싶을 때 | 원본 구조와 비교하여 orientation 변화 분석 |
| back-mutation할 residue를 결정할 때 | 구조적으로 가장 큰 영향을 주는 잔기 확인 가능 |
✅ 컴퓨터 기반 예측 프로그램
어떤 human VH–VL germline 조합이 좋은 orientation을 형성할지 예측할 방법은?
✅ Narayanan et al. 방법
“구조 기반 + 에너지 기반”의 실용적 접근법
핵심 알고리즘
VH–VL interface residue들의 side-chain rotamer들을 다양하게 샘플링함
-> 모든 조합에 대해 에너지 계산 수행
-> Molecular Mechanics (MM) force field를 이용하여 각 조합의 에너지 평가
즉, 어떤 rotamer 조합이 가장 안정적인지를 계산
최적 에너지 조합을 선택
→ 가장 낮은 에너지를 가진 VH–VL 조합 = 구조적으로 가장 안정적일 가능성 높음
backbone (main chain)은 실제 결정 구조에서 가져옴 → 현실 기반 샘플링
🖐 Rotamer
side-chain이 가질 수 있는 여러 입체 배열 형태
✅ Rosetta Antibody의 접근법
"Narayanan 방법과 유사한 방향의 구현체"
핵심 특징
Rosetta는 side-chain 최적화, backbone flexibility, 에너지 기반 scoring을 모두 고려하는 고급 구조 모델링 도구
RosettaAntibody는 VH–VL orientation 예측, homology modeling, humanization 등을 지원
VH–VL interface residue를 중심으로 rotamer 최적화, 구조 스코어링 수행
RosettaAntibody는 CDR loop prediction에도 강점을 가지며
humanization 후 VH–VL orientation이 어떻게 바뀌는지를 비교/평가 가능
✅ VH–VL packing angle 예측 위한 머신러닝과 구조 기반 방법
✅ 머신러닝 기반 VH–VL orientation 예측 방법
567개의 항체 결정 구조에서 VH–VL interface 정보를 수집
Packing angle을 예측하기 위한 머신러닝 모델(신경망)을 학습시킴
중요한 점은:
연구자가 임의로 interface residue를 고르지 않았고,
유전 알고리즘(genetic algorithm)을 사용해 모델 학습에 가장 효과적인 20개 이하의 residue 위치를 자동 선택
핵심 결과:
그 중 13개 residue 위치가 packing angle에 가장 큰 영향을 줌
예측 정확도는 약 ±2도 범위로, 결정 구조에서 측정한 실험적 오차 수준과 비슷함
→ 실용적으로 매우 우수한 정확도
🖐 유전 알고리즘 이란?
VH–VL packing angle 예측에서 가장 중요한 residue 위치 조합을 찾기 위한 최적화 방법
1️⃣ 초기 세대 생성
여러 조합의 residue 위치 리스트(예: 5개, 10개, 20개…)를 무작위로 생성
2️⃣ 각 조합의 성능 평가 (Fitness)
각 조합으로 만든 모델이 packing angle을 얼마나 정확히 예측하는지 평가
예측 오차가 작을수록 높은 점수 (fitness)
3️⃣ 선택 (Selection)
성능이 좋은 조합들만 다음 세대로 선택
4️⃣ 교차와 돌연변이 (Crossover & Mutation)
선택된 조합들을 합쳐서 새로운 조합을 만들거나
일부 residue 위치를 무작위로 바꾸며 다양성 부여
5️⃣ 반복 (진화)
수십~수백 세대 반복하면서
점점 packing angle 예측에 최적화된 residue 조합으로 진화
✅ 또 다른 구조 기반 방법
VH–VL orientation을 5가지 각도(angles) + 1개 거리(distance) 총 6가지 수치로 정량화
각각의 수치를 잘 예측하는 데 영향을 주는 key residue 위치를 식별
결과
총 35개 residue 위치가 orientation 결정에 중요
-> 24개는 VL에서, 11개는 VH에서
단점
35개의 residue를 germline selection에서 모두 고려하기에는 너무 많고 복잡함
→ 실제 humanization 작업에서 적용하기엔 비현실적
✅ Backmutation
생성되는 변이체의 인간성 점수를 감소시키지만, 결합 친화도는 향상될 것
✅ 단순한 CDR grafting의 한계
mouse CDR을 human FR (framework region)에 단순히 붙이면,
VH–VL interface는 잘 유지되더라도, CDR의 입체 구조(conformation)가 무너질 수 있음.
특히, CDR을 지지해주는 특정 FR 잔기들이 달라졌을 때 문제 발생
✅ 해결책: CDR 구조를 유지하는 critical FR 잔기 보존
human germline 선택 시, 단순히 sequence identity가 높은 걸 고르지 말고,
CDR 구조를 유지하는 key FR residue들이 일치하는지 우선적으로 고려해야 함.
→ 일치하지 않는 경우에는 back mutation(즉, human residue를 다시 mouse residue로 되돌림)을 적용해야 함.
✅ Vernier zone의 개념
CDR 구조를 지탱하고 간접적으로 항원 결합에 영향을 주는 FR 잔기 집합
4개는 CDR-H1 일부 (Chothia/Martin/IMGT 정의 기준)
나머지 26개: VH와 VL에 각각 13개씩 분포 (그림 7, 8 참조)
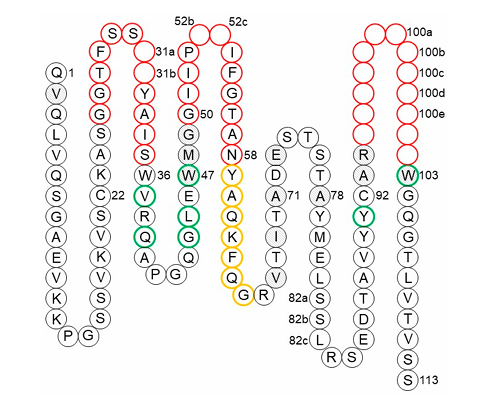
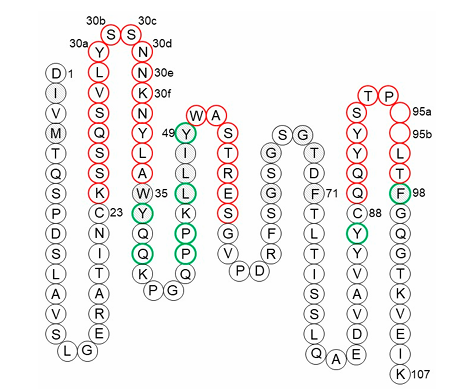
ex)
Heavy chain residue 71번: CDR-H2의 canonical 구조 결정
이 위치를 보존하지 않으면 구조가 무너져 binding 저하됨하지만 모든 항체에서 반드시 중요한 건 아님 (예외 존재)
✅ 실제 권장 전략
CDR 구조 유지에 중요하다고 알려진 FR 위치 추출
항체-항원 복합체 구조 또는 모델링 사용
각 Vernier residue에 대해, 실험적으로 (또는 모델링 기반으로) binding에 영향 주는지 확인
-> 진짜 영향 있을 때만 back-mutation 적용
✅ Deimmunization (탈면역화)
✅ CDR 내부도 인간화할 수 있는가?
일반적으로 CDR은 그대로 유지하고 FR만 인간화하지만,
→ 항원 결합이나 CDR 구조에 영향을 주지 않는 잔기라면,
→ CDR 안에 있는 비인간 잔기도 인간 잔기로 바꿀 수 있음.
🔎 ex)
CD25, VEGF, TNFα를 타겟으로 하는 mouse 항체 3종을 인간화할 때,
반복적인 germline 서열 탐색 + 컴퓨터 알고리즘으로
→ human CDR residue를 최대한 도입해서
→ 결합능 유지 + humanness 향상 성공함.
→ 결과물: daclizumab, bevacizumab, infliximab
✅ CDR이 면역원성의 핵심 부위라는 발견
CD4+ T 세포에 의해 인식되는 면역원성 epitope은 CDR에만 존재하고, FR에는 없음.
따라서 면역반응을 줄이기 위해서는
→ CDR 안의 epitope를 겨냥한 아미노산 변이가 필요함.
→ 이 과정을 deimmunization (탈면역화)라고 부름.
✅ Deimmunization = 면역원성 줄이기 전략
면역원성을 줄이기 위해 T 세포 epitope가 되는 CDR 부위의 아미노산을, 인간 잔기로 1~2개만 바꿔주는 전략.
결합능을 그대로 유지하면서, T 세포 인식 위험을 크게 줄일 수 있음.
✅ 구조 기반 deimmunization Tool: EpiSweep
EpiSweep는 구조적 정보 + 면역예측 알고리즘을 결합해서
→ 면역원성이 높을 수 있는 위치를 예측하고,
→ 결합능과 구조 안정성을 유지하면서 가장 좋은 변이 조합을 제안해 줌.
항체뿐만 아니라 다른 단백질에도 쓸 수 있지만,
→ 항체 deimmunization에 특히 유용함.
✅ Super-Humanization
| 구분 | 설명 |
|---|---|
| 기존 방식 | CDR은 유지, FR은 non-human → human으로 교체, 이때 FR에서 유사한 human 서열을 선택함 |
| Super-Humanization | FR 유사도는 신경 쓰지 않고, CDR의 구조(canonical structure)가 잘 유지될 수 있는 human germline을 선택함 |
즉, CDR 구조 보존에만 집중해서 human FR을 고르는 방식
Super-Humanization의 핵심은?
“CDR 구조가 유지될 수 있는 human germline을 선택해 구조적 호환성만 보장하면, sequence는 달라도 된다”
→ FR과 CDR이 조화롭게 작동하는 구조적 스캐폴드를 만드는 것이 목적
But 보통 성능 감소가 크기 때문에, 일반적으로는 많이 사용되지 않음
✅ Humanness Optimization
✅ Humanness란 무엇인가?
항체 서열이 얼마나 인간 항체와 유사한가를 나타내는 개념
면역원성(immunogenicity)을 줄이기 위해, 항체가 인간 항체처럼 보이도록 함
1️⃣ Human String Content (HSC) Score
실험적으로도 검증됨
| 항목 | 설명 |
|---|---|
| 정의 | 특정 항체 서열 내에 인간 germline과 일치하는 아미노산 비율 |
| 계산 방법 | target 항체의 각 위치에 대해 가장 유사한 human germline 서열과 정렬하고, 일치하는 잔기 개수를 세어 비율 계산 |
| 특징 | - 국소적(local) 정보 기반 - 단일 germline이 아니라 여러 germline에서 유사한 패턴들을 모아서 평가 |
| 장점 | - 단순한 전체 identity보다 면역원성과 더 밀접한 상관관계 - 다양한 humanized 후보군을 만들 때 유용 |
2️⃣ ACE (Analogous Contact Environments) 필터
구조적 적합성 체크
| 항목 | 설명 |
|---|---|
| 문제 | HSC 기반으로 여러 germline 조각을 짜맞추다 보면 구조 충돌(clash) 위험 있음 |
| 해결 | ACE 필터: 다양한 antibody 구조 데이터베이스에서 유사한 잔기 조합과 구조 환경이 있는지 확인 |
| 방식 | 구조적으로 가까운 잔기 간의 조합을 precedence score로 평가함 (높을수록 안정적인 구조 패턴) |
3️⃣ 다른 humanness 스코어링 방법들
| 스코어링 방식 | 설명 |
|---|---|
| 평균 유사도 | 전체 germline DB와 pairwise identity 평균 |
| Top-20 평균 | 가장 유사한 20개 germline 서열의 평균 |
| 9-mer window | 서열을 9-residue 단위로 끊고 최고 유사도 측정 |
| pairwise dependency | 특정 위치 간의 pair correlation도 고려 → 이는 CDR/FR을 구분하지 않는 방식 FR에도 항원 결합에 중요한 잔기가 있음을 반영한 방식 |
✅ Humanness score VS 실제 면역원성
여러 연구에서 높은 humanness score가 실제로 면역원성 감소와 상관관계 있음을 확인
임상데이터 기반으로도 면역반응이 덜 일어나는 항체일수록 humanness score가 높음
✅ Affinity Maturation
✅ 핵심 목표: 항체의 결합 친화도(affinity) 를 향상시키는 것
항체가 항원과 더 강하게 오래 결합하도록 만들어 치료 효과를 높이는 것
핵심 메커니즘: 결합 해리속도(k_off) 를 낮춰서 전체 KD를 줄이는 방식
✅ 주요 사례 1: Anti-integrin antibody VLA1
PDB 기반 crystal 구조가 있다면, interface residue를 직접 확인하고 돌연변이 후보를 도출 가능
1️⃣ side-chain packing 향상: 공간적으로 잘 맞도록 잔기 교체
결정 구조(crystal structure) 를 참고하여 항원 접촉 잔기 전반에 대해 >80개 단일 변이(mutations) 설계
2️⃣ electrostatic interaction 최적화: 전하 상호작용 강화
항체–항원 결합면의 전하 분포(Electrostatics)를 계산적으로 분석
"이 위치에 이 전하성 아미노산을 넣으면 결합력이 증가할 것이다"를 예측
이 두개의 접근법을 적용했을 때, 실제로 affinity 10배 증가
✅ 주요 사례 2: trastuzumab / rituximab
중심 접촉 잔기(mutating core paratope residues) 방식 대신, 주변(peripheral) 잔기를 타깃으로 하는 새로운 접근법
| 구분 | 기존 방식 | 새로운 전략 |
|---|---|---|
| 타겟 위치 | 항원-contact 중심 잔기 (core paratope) | 인터페이스 주변 잔기 (periphery) |
| 문제점 | 이미 tightly packed되어 있고 최적화돼 있음 → 더 이상 크게 개선 안 됨 | hydrophilic하고 solvent-exposed → 전하 상호작용(Electrostatics) 향상 가능 |
| 기대효과 | 결합력 증가 | long-range interaction 향상, 예기치 못한 유익한 변이 가능 |
✅ In silico 기반 Affinity Maturation 설계
1️⃣ 단순 mutation
60개 CDR 잔기 전체에 대해 각 잔기에 대해 20가지 아미노산으로 단일 치환 돌연변이 생성
에너지 기반 스코어링: Monte Carlo + molecular dynamics 사용
-> 총 자유에너지(binding free energy) 로 돌연변이의 효과를 정량화
상위 11개 변이 실험적으로 테스트 -> 그 중 2개만 실제 친화도 향상
2️⃣ 대안 전략: in vivo 빈도 높은 아미노산만 사용
항체가 체내에서 성숙(affinity maturation)되는 과정인 somatic hypermutation(SHM)에서 자주 등장하는 아미노산만을 후보로 제한하여 돌연변이 설계
-> 돌연변이 수는 줄지만, 친화도 향상 성공률이 60~70%로 대폭 향상
🖐 Somatic Hypermutation (SHM)란?
B세포가 항원을 인식한 후, 그 B세포의 항체 유전자(V(D)J 영역)에 의도적으로 돌연변이를 일으키는 과정
→ 돌연변이된 항체 중 일부는 더 강하게 항원에 결합하게 됨
→ 결과적으로 항체의 친화도(affinity)가 점점 높아짐
→ 이 과정을 "affinity maturation"이라고 함
🖐 어떤 유전자에서 일어나는가?
SHM은 가변 영역(variable region)—즉, V(D)J 재조합으로 생성된 영역에서 발생
특히 CDR (Complementarity-Determining Region)에서 돌연변이가 집중됨
→ 항원 결합부위니까 친화도 변화에 직접적 영향
🖐 어떻게 돌연변이가 일어나는가?
Activation-Induced Cytidine Deaminase (AID)
→ AID가 DNA의 C를 U로 바꾸는 돌연변이 반응을 촉진
→ 이후 DNA 수선 과정에서 무작위적인 염기 치환(mutation)이 발생
🖐 Activation-Induced Cytidine Deaminase (AID) 란?
B세포에 특이적으로 발현되는 효소로,
DNA의 시토신(Cytidine)을 우라실(Uracil)로 탈아민화(deamination)해서 돌연변이를 유도함
AID는 면역계의 편집기(editor)
→ 항체 유전자를 고의로 ‘오타’내서
→ 그중 우연히 더 강한 항체가 만들어지면 선택되는 구조 (진화 시뮬레이션)
이상한 U가 생겼으므로, DNA 수선 시스템이 작동
->
다양한 방법으로 수선되며 염기 치환(point mutation) 또는 이중 가닥 절단(double-strand break) 발생
그 결과:
- SHM → 다양하고 친화도 높은 항체 생성
- CSR → 동일 항원에 대한 다양한 기능의 항체 생성 (예: IgG는 opsonization, IgE는 알레르기 반응 등)
✅ 중요한 발견: "친화도 향상 변이는 SHM hotspot에 집중된다"
체내 somatic hypermutation(SHM)은 무작위가 아니라,
특정 뉴클레오타이드 패턴에 선호적으로 일어남
대표 hotspot motif
-> RGYW / WRCY / TAA
이들 부위는 CDR과 FR 사이 경계, 또는 interface 주변부에 자주 존재
실제 실험에서 확인된 바:
친화도 증가 돌연변이들이 이러한 SHM hotspot 근처에 위치하는 경우가 많음
→ in silico 설계 시에도 이런 germline hotspot 기반 정보 활용이 효과적
🖐 SHM hotspot motif (RGYW / WRCY / TAA)
AID 효소가 돌연변이를 잘 일으키는 특정 DNA 염기서열 패턴
1️⃣ RGYW / WRCY란?
IUPAC 코드로 표현된 뉴클레오타이드 서열 패턴
(유전자의 특정 염기 조합에 대해 돌연변이가 잘 일어나는 곳)
2️⃣ WRCY는 RGYW의 반대 가닥 (reverse complement)
실제 DNA는 이중가닥(double-stranded)이라, 한 가닥에 RGYW가 있다면 반대 가닥엔 WRCY가 있음
→ 그래서 둘 다 hotspot으로 간주됨
| 코드 | 의미 |
|---|---|
| R | Purine (A or G) |
| Y | Pyrimidine (C or T) |
| W | Weak (A or T) |
| S | Strong (C or G) |
🔎 ex) RGYW
R - G - Y - W가능한 조합
A-G-C-A
A-G-T-A
G-G-C-T
G-G-T-T
등
→ 이런 4개 염기 조합이 DNA에 있으면 AID가 잘 작동해서 SHM이 잘 일어남
3️⃣ TAA motif
TAA는 AID가 선호하는 단순한 3-mer motif 중 하나
특히 translation stop codon이기도 해서
→ SHM 중에 부작용으로 조기 종료 돌연변이가 생길 수도 있음
→ 이 경우 해당 B세포는 사멸됨 (negative selection)
🖐 hydrophilic (친수성)
물을 좋아하는 성질을 가진 분자 또는 잔기
전하를 띠거나 극성(polar)을 띠는 아미노산 잔기들이 여기에 속함
대표적 친수성 아미노산: Lys, Arg, Asp, Glu, Asn, Gln, Ser, Thr, His 등
- 단백질 표면이나 용매에 노출된 위치에 자주 존재
- 물(H₂O) 분자와 수소결합을 잘 형성함
- 주로 항체-항원 인터페이스 주변부(periphery) 에 위치
- electrostatic interaction 향상에 기여 가능
🖐 solvent-exposed (용매 노출)
단백질의 특정 아미노산 잔기가 용매(보통 물)에 노출되어 있는 상태
-
보통 hydrophilic residue가 solvent-exposed됨
-
결합 인터페이스 주변, CDR 바깥, loop 영역에 자주 존재
-
구조적으로 접근성(accessibility)이 높아, 돌연변이에 쉽게 대응 가능
-> 이런 위치는 돌연변이 도입 시 안정성이나 구조를 크게 해치지 않음
-> affinity maturation이나 engineering에 안전한 타깃
✅ 3D 모델링 기반 affinity maturation 전략
1️⃣ 항체–항원 복합체의 3D 모델은 변이 설계의 핵심
항체 친화도 향상(affinity improvement)을 위해서는 항체-항원 결합 부위의 3차원 구조 정보가 필수.
실험 구조가 없다면, computational docking(결합 예측)을 통해 복합체 모델을 생성할 수 있음.
2️⃣ Docking 정확도와 한계
항원 단독 구조만으로 항체 결합 복합체 모델을 생성
일부 정확한 도킹 예측도 가능했으나, 올바른 결과와 잘못된 결과를 구분하는 데 어려움이 있음
→ Scoring function 개선이 중요
🔎 ex) anti-gastrin 항체 TA4
항원: 7-residue linear peptide (gastrin)
항체: scFv TA4
목표: 실험 구조 없이 친화도 454배 향상📌 접근법
항체와 항원을 도킹해 복합체 3D 모델 생성항체-항원 복합체의 구조 모델은 선형 에피토프를 나타내는
7개 residue 펩타이드를 항체 모델에 도킹하여 생성Molecular dynamics로 결합 자세 정제
→ 펩타이드가 α-helix 구조를 갖는다는 점 확인CDR-H3 4개 잔기와 CDR-L3 5개 잔기를 randomization 대상으로 선정
단계적 library 설계 및 스크리닝 수행:
1차 라이브러리 (CDR-H3): 10배 향상된 double mutant
2차 라이브러리 (CDR-L3): 추가 2배 향상
3차 설계: 나머지 CDRs에 확장 → 수 nM 수준 친화도 도달
✅ 핵심: 구조 기반 모델만으로도 실험적 구조 없이도 정확한 위치 선정
→ 효과적인 라이브러리 설계 → 친화도 대폭 향상
🖐 왜 항원 전체 대신 에피토프 펩타이드만 사용하는가?
1️⃣ 계산 자원과 시간 절약
항원 전체 구조는 수백~수천 개의 아미노산을 포함하며, 구조적으로 유연한 영역도 많음
이렇게 큰 분자 전체를 도킹에 사용하면 도킹 후보군이 너무 많아지고 계산 비용이 급증
반면, 실제 항체와 상호작용하는 에피토프는 보통 6~15개의 잔기 정도이므로,
짧은 펩타이드만 추출하여 도킹하면 계산이 빠르고 정확도도 향상
2️⃣ 정확한 결합 위치 예측 가능
항체는 항원 전체와 결합하지 않고, 특정 표면 패치(=선형 또는 구조적 에피토프)와만 상호작용
전체 항원을 쓰면, 도킹 프로그램이 엉뚱한 위치(비에피토프 영역)에 결합하도록 예측할 위험이 있음
따라서 결합 부위만 남기고 나머지는 제거함으로써 도킹 정확도 상승 가능
3️⃣ 구조 변형과 유연성 문제 회피
항원 전체는 구조적으로 복잡하고, 여러 유연한 loop 영역을 가질 수 있음.
에피토프 부분만 따로 도킹하면, 그 부위만의 정확한 상호작용을 모델링하는 데 집중할 수 있음.
특히 선형 epitope (linear epitope)인 경우, 짧은 펩타이드로 독립적 도킹이 매우 효율적임.
✅ CDR 중심의 라이브러리 디자인과 구조 기반 affinity maturation 전략
1️⃣ 항체–항원 결합에 가장 많이 관여하는 central CDR들(H3, L3)을 중심으로 library 설계가 많이 이루어짐.
최적화된 VL을 설계 -> 최종적으로 VH와 조합
2️⃣ CDR-H3가 너무 중요할 경우, H2 또는 L1, H1도 타깃
때때로 H3가 이미 매우 강하게 항원과 결합 중이면,
H2 또는 다른 CDR들(L1, H1 등)로 diversification 범위 조절.
3️⃣ CDR 외 영역도 조절 가능: domain core 또는 elbow region
단순히 CDR이나 Vernier zone 잔기만이 아니라, V–C 도메인 연결 부위(elbow region) 등의 구조적 유연성도 중요.
🔎 ex) metelimumab (anti-TGF-β1 항체)
scFv → IgG4로 변환 시 결합력이 50배 낮아짐.
원인 추정: IgG 구조의 유연성 감소.해결책: heavy, light chain의 elbow 부위에 glycine 2개 삽입 → 유연성 증가
→ 결합 친화도 복구 + CDR 간 거리 확장 → 항원과 새로운 결합 형성
✅ Specificity
| 개념 | 의미 및 왜 중요한가 |
|---|---|
| Epitope-specific binding | 항체는 특정 항원을 선택적으로 인식함 → 특이성이 좋을수록 부작용↓ |
| Cross-reactivity | 다른 비슷한 항원에도 붙을 수 있음 (문제점) |
| 멀티스페시픽 항체 | 유사 항원에 교차 결합 → 의도치 않은 타깃 공격 위험 |
| 특이성 조절 방법 | CDR-H3, H1, H2, L3 등을 중심으로 디자인 + 에너지 기반 스코어링 + 구조 기반 필터링 |
✅ CMC Considerations
| 개념 | 설명 |
|---|---|
| PTMs (후천적 변형) | 아미노산이 산화/탈아미드화/글라이케이션 되며 구조/기능이 바뀜 |
| Deamidation, Oxidation | 항체의 안정성, 효능, PK에 악영향. 주로 Asn, Met 등에서 발생 |
| Glycosylation | 당이 붙는 정도나 위치가 약물 효능/반감기에 큰 영향 |
| 클리핑/응집 | 단백질 분해 혹은 비정상 응집 발생 시 품질 저하 |
CDR/FR에 PTM hotspot (예: Asn-Gly, Met, Trp 등) 있는지 확인 → 위험도 부여
Stability 예측 score에 산화/탈아미드화 위험 가중치 추가
Glycosylation 부위 존재 여부 → 생산성과 PK 예측에 반영
✅ Solubility
문제: 고농도 항체 제형(>100 mg/mL)에서 점도(viscosity) ↑ or 응집(aggregation) ↑
→ 피하주사(s.c. injection)가 불가능해짐 → 약물로 개발 불가
| 전략 | 적용 방식 | 사례 |
|---|---|---|
| ① Hydrophobic patch 제거 | VH–VL 사이 등 표면 소수성 영역에 친수성 변이 | V45K → solubility↑, binding 유지 E60Y → electrostatic neutralization |
| ② 전하 조절 | 음전하 영역 중화/변형 → 점도 감소 | E60Y vs E60K: 후자는 오히려 destabilizing |
| ③ CDR 영역 내 hydrophobic cluster 해체 | 특히 CDR-H3, L3의 소수성 잔기에 전하 부여 | Alzheimer 단일 VH 항체: CDR3 양쪽에 음전하 삽입해 aggregation 억제 |
| ④ 글리코실화 도입 (Glycosylation) | Aggregation hotspot 덮기 위해 CDR에 glycan site 삽입 | CNTO607에서 D53N 변이로 solubility↑, affinity 유지 |
| ⑤ pI 조절 | 결합 부위와 무관한 위치에 전하 잔기 삽입해 등전점 조정 | CNTO607: pI 변경으로 용해도 2배 향상 |
| ⑥ 컴퓨터 모델링 기반 스크리닝 | Fv domain 모델링 → hydrophobic patch 탐지 후 변이 시뮬레이션 | integrin 항체에서 97개 변이 예측, solubility↑ 유지결합 |
| ⑦ Aggregation 예측 알고리즘 | AggScore: 소수성+전하 패치 분포 기반 집합 가능성 예측 | Adnectin 학습, 실제 임상 mAb에도 적용됨 |
✅ Stability
✅ 왜 Stability가 중요한가?
항체가 잘 만들어져도 불안정하면 약으로 개발할 수 없음
→ 저장 중 변성, 응집, 분해 등 문제 발생
특히 CDR loop가 너무 유연하거나, 프레임워크에 취약한 위치가 있으면 구조가 무너지기 쉬움
✅ Stability에 영향을 주는 주요 요소
| 요소 | 설명 |
|---|---|
| 1. Core packing | 도메인 내부 소수성 코어가 잘 쌓여 있지 않으면 열에 약함 |
| 2. VH–VL 인터페이스 상호작용 | VH와 VL 간 결합력이 약하면 전체 구조 불안정 |
| 3. 내부에 묻힌 전하/극성 잔기 | 내부에 bury된 polar residue → 물 없이 불안정함 |
| 4. 수소결합 네트워크 | 잘 짜인 hydrogen bond network → 안정성 ↑ |
| 5. 표면 전하/극성 분포 | 무질서한 표면 전하분포 → 응집↑, 변성↑ 가능 |
이런 요소들을 고려해서 불안정하게 보이는 잔기들을 찾고, 돌연변이(변이체) 설계를 통해 Tm (열 안정성)을 향상시킴
Tm (Thermal Transition Midpoint) 값이 높을수록 안정한 항체
✅ Antibody Activity Engineering
✅ Binding Domain Engineering
1️⃣ Single Domain Antibodies (sdAbs, VHH/nanobody)
VHH (Camelid nanobody)의 특성
-
무게: 약 12–15 kDa로 매우 작음.
-
출처: 낙타류(Camelid)에서 유래된 heavy chain-only antibody에서 파생된 단일 도메인 항체.
-
구조: 일반적인 IgG와 달리 light chain이 없음.
따라서 더 친수성(hydrophilic)이며, 용해성(solubility) 높고, 응집(aggregation) 적음. -
장점
작은 크기 덕분에 GPCR처럼 접근이 어려운 에피토프도 타겟팅 가능.
CDR-H3가 더 길어 구조적으로 깊은 포켓에도 결합 가능.
약물 투여 경로의 다양화 가능(주사, 흡입, 경구, 점안) -
인간화 가능성
VHH 서열을 인간의 IGKV/IGLV와 정렬 가능함 (canonical 구조 유사성).
따라서 면역원성(ADA)을 줄이기 위한 인간화(humanization)가 가능.
2️⃣ VNAR (상어 유래 단일 도메인 항체)
VNAR의 특성
-
출처: 연골어류
-
크기: 11–14 kDa로 VHH보다 더 작음.
-
구조적 특징:
8개의 β-sheet (VHH나 IgG는 10개)
CDR2 없음, 대부분의 다양성은 CDR3에서 유래
긴 hinge region 없음 → 구조가 단순함 -
장점
열적 안정성 매우 우수, 다양한 시스템에서 고수율로 생산 가능.
좁고 독특한 결합 부위(niche)에 잘 들어갈 수 있음.
CXCR4, HER2, PD-1, glypican-3 등 다양한 질환 표적에 대한 binder 개발 완료.
✅ Avidity
항체는 두 개의 Fab 항원결합 부위를 가지며,
표적 세포 표면에 여러 항원(epitope)이 있을 경우 두 Fab이 동시에 결합 가능
→ Multivalent binding (다가성 결합).
이때 발생하는 누적 결합력을 Avidity라 함
✅ Avidity 효과 발생 조건
- 항원이 충분히 밀집되어 있어야 함.
- 첫 Fab이 결합한 상태에서 두 번째 Fab이 결합할 기회가 있어야 함.
Fab–epitope 결합의 공간적 제약(stereic occlusion) 때문에,
→ 두 Fab이 동시에 결합하기 어렵거나 못할 수 있음.
이런 경우, avidity 효과가 제한됨 → 중화(neutralization) 능력 감소.
🔎 ex)
항원이 세포 표면에 너무 멀리 떨어져 있거나,
결합할 구조가 공간적으로 좁아서 두 Fab이 같이 결합 못하는 경우
✅ FcγR을 통한 Avidity 증가
🖐FcγR(Fc 감마 수용체) 란?
면역세포(대식세포, NK세포, 수지상세포 등)에 존재하는 수용체
항체의 Fc 영역(꼬리 부분)과 결합하는 수용체 (항체가 Fc 부분으로 신호를 전달하는 관문)
→ 항체가 항원을 인식하고, Fc 부분이 FcγR과 결합하면
→ 면역세포는 그 신호를 받아서 공격 시작
항체의 Fc 부위가 FcγR(면역세포 수용체)에 결합하면,
항체들이 면역세포 표면에 클러스터링됨.
결과적으로 여러 Fab이 가까이 있게 되어 항원에 대한 다가 결합력(Avidity)이 간접적으로 증가함.
🖐 Effector function (면역세포의 작용)
항체가 FcγR과 결합했을 때 면역세포가 활성화되며 수행하는 역할들.
- ADCC: NK 세포가 표적 세포를 죽임
- Phagocytosis: 대식세포가 표적을 잡아먹음
- Cytokine 분비: 염증 유도
✅ FcγR 결합의 양면성
| 긍정적 효과 | 부작용 위험 |
|---|---|
| Fab Avidity ↑ → 타겟 결합력 ↑ | 면역세포 과활성화(=effector function) → 자가면역 위험 |
| ADCC, 식작용, 염증 반응 유도 가능 | 정상세포 손상 가능 (예: SLE, 루푸스 등) |
이를 억제하기 위해 IVIG(고용량 면역글로불린)를 사용해 FcγR 차단
→ 하지만 부작용(염증, 혈전 등) 위험 있음.
✅ 항암 면역조절 항체의 Fc 조절 전략
🖐 면역관문 억제제 (immune checkpoint inhibitor)
PD-1, CTLA-4는 T세포의 "브레이크 역할"을 하는 단백질
이를 억제하면 T세포가 다시 활성화되어 암세포를 공격하게 됨
→ 그래서 항체로 PD-1 또는 CTLA-4를 차단함
1️⃣ 면역관문 억제제 (PD-1, CTLA-4) 를 target
면역관문 수용체를 타겟
-> T세포의 브레이크 역할을 차단
→ T세포를 재활성화시켜 종양 공격 유도
이때 Fc 부위는 silent하게 설계 (FcγR 및 보체 C1q 결합 제거)
-> T세포를 죽이는 불필요한 ADCC/CDC 유도 방지 위해
✅ 왜 이걸 막아야 할까?
PD-1이나 CTLA-4는 암세포가 아니라, 우리 몸의 T세포 표면에 있음
항체가 PD-1을 인식하면 T세포에 붙음
이 상태에서 Fc가 활성화되면 (Fc effector 기능이 작동하면) ?
→ T세포가 ADCC나 CDC로 죽어버림
→ T세포는 암세포를 죽여야 하는데, 오히려 자기 몸이 죽는 역효과 발생
→ 그래서 "Fc silent"하게 설계 (FcγR에 잘 결합하지 않도록 돌연변이 삽입)
→ 항체는 T세포의 브레이크만 해제하고, 불필요한 면역세포 작동은 유도하지 않음
→ 더 안전하고 효과적인 치료
🖐 ADCC (항체의존 세포독성)
NK 세포가 항체 붙은 세포를 죽임
🖐 CDC (보체 의존성 세포독성)
보체계가 활성화되어 세포를 용해시킴
✅ 면역자극 수용체 타겟 항체 (agonist antibody)
보통은 Fc effector 기능을 억제(silent Fc)시키지만,
OX40, GITR 항체는 일부러 effector 기능을 남겨서 Treg 세포를 죽이는 데 활용함.
→ 활성화 보다 제거 가 항암 효과 핵심일 수도 있다는 전략
Target: TNF receptor 계열 (OX40, CD28, 4-1BB 등)
→ 면역 자극 수용체인 T세포나 APC 표면에 존재, T세포를 활성화·증식시킴
PD-1/CTLA-4 억제 항체는 단순 억제뿐 아니라,
T세포 또는 항원제시세포(APC)에 있는 면역자극성 수용체를 타겟할 수도 있음
→ T세포 활성화 및 증식 유도
→ 강한 항암 면역반응
→ OX40, GITR을 타겟하는 특별한 agonist 항체
OX40, GITR: T세포 표면에 존재하는 자극 수용체
👉 보통 T세포를 활성화시키는 역할
그런데 종양 조직 안에서는, 조절 T세포(Treg)도 OX40, GITR을 발현
👉 Treg는 면역을 억제해서 종양이 도망갈 수 있게 만듦.
따라서 OX40/GITR 항체가 이 수용체에 결합하고,
그 Fc 영역이 FcγR에 결합해서 ADCC/ADCP (세포 사멸 유도)를 일으키면
👉 Treg를 선택적으로 제거할 수 있음
| 항목 | 설명 |
|---|---|
| OX40, GITR | T세포에 있는 면역 자극 수용체 (TNFR 계열) |
| 문제 | 종양 미세환경에선 Treg도 이 수용체를 가짐 → 면역 억제 유지 |
| 전략 | 항체가 이 수용체에 붙고, Fc가 면역세포 FcγR에 붙어 → Treg를 죽이게 함 |
| 이유 | Treg를 제거하면 전체 항암 면역이 더 잘 활성화됨 |
| 결과 | 단순히 effector T세포를 자극하는 것보다, Treg 제거가 항암에 더 효과적일 수도 있음 |
🖐 Agonist (작동자)
수용체를 활성화시켜 면역반응을 증폭
🖐 면역효과세포(T세포, NK세포)
TNF 수용체 계열의 활성화 수용체가 존재
🛑 But
실제로 "정상적인 T세포"도 죽을 위험이 존재
그래서 이 전략은 종양 내부에서만 선택적으로 작용하게 설계해야 하고,
그게 굉장히 중요한 항암항체 개발 포인트
1️⃣ OX40, GITR 같은 TNFR 계열 수용체는 모든 T세포에 있을 수 있음
- Effector T세포 (암 공격하는 애들)
- Regulatory T세포 (Treg) (면역 억제하는 애들)
둘 다에 발현될 수 있음
👉 그래서 항체가 무작정 이 수용체를 인식하면 정상 T세포도 공격당할 수 있음.
2️⃣ 하지만 실제로는 종양 미세환경(TME)에서만 작동하게 유도함
Fc effector 기능이 종양 내 Treg 제거에만 작용하도록 조정
종양 내 FcγR 발현 환경, Treg 밀도, 항체 투여량/타겟 특이성 등을 고려해서
👉 비종양 조직의 정상 T세포는 되도록 안 건드리게 설계
✅ Antibody–Drug Conjugates, ADCs
✅ ADC의 기본 개념
항체의 특이성 + 소분자 독성약물의 강력한 세포사멸 능력을 결합한 치료제.
타겟 항원만 발현하는 암세포에만 약물을 전달함으로써, 정상 세포에는 독성 최소화.
✅ 구성 요소
| 구성 요소 | 역할 |
|---|---|
| 항체 (Ab) | 종양세포 특이 항원 인식 및 결합 |
| 링커 (Linker) | 약물을 항체에 연결해주는 화학적 연결 고리 |
| 약물 (Payload) | 암세포를 죽이는 고독성 화합물 |
✅ Fc Activity Engineering
1️⃣ 항체는 항원 결합(Fab) 만으로는 충분하지 않다
항체가 표적에 결합해도 Fc 영역이 면역 세포를 잘 활성화하지 않으면 효과가 떨어짐.
2️⃣ Fc 영역의 avidity (발현 방식, 배열 등)도 중요하다
Fc가 어떻게 배열되어 있는지 (즉, Fc 수용체에 얼마나 잘 보이는지)가 면역세포 활성화에 큰 영향.
FcR (ex. FcγRIIIa)이 인식 잘 해야 NK 세포, 대식세포가 활성화됨
→ ADCC 등 유도 가능.
3️⃣ Fc 조작 (Fc Engineering)을 통해 면역 활성 조절 가능
Fc 영역을 유전공학적으로 조작하여,
면역 반응(효능)을 높일 수 있음
약물의 물성 (안정성, 반감기 등)을 향상시킬 수 있음
🔎 ex 1) IgG subtype 변경 (ex. IgG1 ↔ IgG2 ↔ IgG4)
IgG1: ADCC 강함
IgG2/4: ADCC 약하거나 없음 → 부작용 줄이기
🔎 ex 2) Fc point mutation
특정 아미노산을 교체해 FcR 결합력 강화/약화
Ex) S239D/I332E → ADCC 증가
4️⃣ Fab과 Fc는 상호영향을 줄 수 있다
Fab이 항원을 결합하면 항체 구조 전체에 영향을 줘서 Fc 기능도 바뀔 수 있음 (구조적 allostery).
즉, Fab–Fc는 독립적이지 않고 상호작용함 → Fc 개량만 따로 평가하는 건 위험할 수 있음.
✅ Fc 영역을 변형시켜 effector function (ADCC, CDC 등)을 조절하는 다양한 전략
항체의 Fc 영역은 면역세포 활성화(ADCC, ADCP, CDC 등)에 핵심적 역할을 하며,
이를 조작함으로써 항체의 작용을 극대화하거나 최소화할 수 있음
| 전략 | 조절 가능한 기능 |
|---|---|
| Fc 아미노산 변이 | ADCC, CDC 증진 or 억제 |
| Fc 당쇄 조작 | ADCC 조절, FcR 결합력 변경 |
| Fc 서브타입 변경 | 면역 반응 강도 조절 |
| Fc 완전 무력화(silent) | 부작용 방지 (특히 면역세포 타깃 시) |
1️⃣ Fc 아미노산 변이에 의한 조절
FcγRIIIa와의 결합력을 높여 NK 세포 활성 → ADCC 증가
FcγR과의 결합을 억제하여 불필요한 면역 반응을 방지
보체(Complement) 활성화 강화 → CDC 유도
2️⃣ Fc 당쇄(Glycosylation) 조절
핵심 위치: Asn297
이곳의 N-linked glycan이 FcR 인식 및 ADCC 조절에 핵심
당쇄 제거 (→ 기능 저해) -> FcR과 결합 불가 → ADCC, CDC 감소
당쇄 최적화 (→ 기능 강화) -> ADCC 대폭 증가 (FcγRIIIa 결합력 증가)
3️⃣ 완전한 Fc 비활성화 전략
특정 상황 (ex. 면역세포를 표적으로 할 때)에는 Fc 기능이 아예 없어야 안전
Fc 무력화 (Fc silent) 전략 채택
✅ Fc region 변이를 통해 항체의 반감기를 조절하는 방법
✅ FcRn과 항체의 반감기
항체의 반감기(half-life)를 FcRn (neonatal Fc receptor)이라는 수용체를 통해 조절
🖐 FcRn이란 무엇인가?
항체(IgG)를 재활용해서 반감기를 연장시키는 수용체
항체를 엔도좀에서 분해되지 않게 보호하고, 다시 혈액으로 되돌림
| 환경 | pH | FcRn–IgG 결합력 |
|---|---|---|
| 엔도좀 내부 | 약산성 (pH < 6.5) | 🔒 강하게 결합 |
| 세포 외 혈액 | 중성 (pH ≈ 7.4) | 🔓 약하게 결합 → 혈액(pH 7.4)에서 방출됨 |
🖐 작동 과정
1️⃣ 혈액의 IgG가 세포에 흡수됨 (비특이적 핀사이토시스)
2️⃣ 엔도좀(산성 환경)에서 FcRn이 IgG에 강하게 붙음 → 분해 안 됨
3️⃣ FcRn–IgG 복합체가 세포 바깥(혈액)으로 이동
4️⃣ pH 7.4에서는 결합력이 낮아져서 IgG를 놓음 → 다시 혈중으로 돌아감
5️⃣ IgG가 분해되지 않고 오래 살아남음 → 반감기 ↑
✅ Fc 변이를 통해 반감기 조절하는 이유
항체를 오래 유지하고 싶다면?
→ FcRn에 잘 붙어야 함 (pH 6.0에서)
→ 하지만 pH 7.4에서는 쉽게 떨어져야 함
| 경우 | 결과 |
|---|---|
| pH 6.0에서 결합 ↑, pH 7.4에서 잘 떨어짐 | 항체가 재활용 → 반감기 증가 |
| pH 6.0과 7.4에서 둘 다 잘 붙음 | FcRn이 항체를 혈중에서 계속 붙잡음 → 재활용 실패 → 분해됨 |
✅ Bispecific Antibodies (BsAbs)
✅ 왜 이중항체(BsAb)를 쓰는가?
1️⃣ 단일항체(mono-specific Ab)의 한계
하나의 항원을 타겟으로 하는 항체는 내성(resistance) 발생 가능성 있음.
신호 전달 경로(crosstalk)로 인해 하나만 막아서는 효과 부족
일부 환자에겐 효과가 없거나 시간이 지나면 무효화됨.
🖐 신호 전달 경로(crosstalk)란?
서로 다른 세포 내 신호 전달 경로들이 서로 영향을 주고받는 현상
2️⃣ BsAb의 장점:
| 특징 | 설명 |
|---|---|
| Dual targeting | 두 항원 또는 epitope을 동시에 인식 가능 |
| 신호 억제 시너지 | ex) EGFR + MET 같이 타겟 → 종양 회피 경로 봉쇄 |
| T세포 리디렉션 | ex) CD3 × 종양항원 BsAb → T세포가 직접 종양세포 공격 |
| 제형 간소화 | 단일 제형이라서 승인·생산·관리 용이 |
| PK 개선 가능성 | Fc 포함 시 → FcRn 결합 통한 반감기 조절 가능 |
🖐 신호 억제 시너지 (Dual signaling blockade)
암세포가 사용하는 두 개의 생존/성장 경로를 동시에 차단해서
회피(crosstalk)나 내성(resistance)을 방지하는 전략
| 항원 | 기능 |
|---|---|
| EGFR | 종양 세포의 성장·분열 신호 전달 |
| MET | 암세포 침습, 이동성, 생존성 유지에 관여 |
EGFR만 차단하면 → 암세포가 MET 경로로 우회
MET만 차단해도 → EGFR이 대체 경로 활성화
EGFR과 MET을 동시에 타겟하는 이중항체(BsAb)를 사용하면,
→ 상호 보완적으로 억제 → 암세포 생존성 대폭 감소
🖐 T세포 리디렉션 (T cell redirection)
T세포를 종양세포 표면으로 직접 유도해서
T세포가 종양을 공격하게 만드는 이중항체 전략
| 항원 | 설명 |
|---|---|
| CD3 | T세포의 활성화 신호 전달 부위 (모든 T세포가 가짐) |
| 종양항원 (ex. CD19, HER2) | 종양세포에 특이적으로 발현됨 |
작동 방식
1. BsAb의 한 팔은 종양세포의 항원에 결합
2. 다른 팔은 T세포의 CD3에 결합
3. BsAb가 종양세포와 T세포를 물리적으로 연결
4. T세포가 종양세포에 면역 시냅스 형성 → 살해 작용 유도
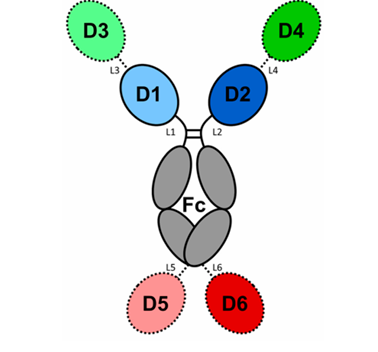
| 구성 요소 | 기능 설명 |
|---|---|
| D1–D6 | 다양한 항원 결합 도메인 (Fab, scFv, nanobody 등) |
| Fc | IgG Fc 영역 → FcRn 결합(반감기 증가), FcγR 결합(ADCC 등 유도) |
| L1–L6 | Linker → 도메인 간 유연성 및 거리 조절 |
| 확장 도메인(D3–D6) | 3개 이상 항원도 타겟 가능 → Multispecificity 구현 |
| Fc engineering 가능 | albumin binding, PEGylation 등으로 PK(약물동태학) 조절 가능 |
기본 항체: D1 = D2 (Fab pair), L1 = L2, Fc = IgG1 Fc
✅ Bispecific Fragments (이중특이성 항체 단편)
항체의 Fab 또는 scFv 단편만을 활용해, 크기를 줄이고 조직 침투력을 높이며,
특정 effector 기능 (ex. T 세포 유도)을 추가하기 위한 다양한 단편 기반 이중특이성 포맷
Fc 없는 bsAb 단편은 작고 침투력 높아 종양 타깃에 적합
다양한 설계 포맷들이 있고, 크기/구성/정제/기능성 등에 따라 장단점 존재
🖐 scFv란?
VH + VL을 짧은 linker로 연결한 최소 항원 결합 단위 (~25 kDa)
✅ IgG-like bispecific antibody (이중특이성 항체)
두 종류의 항원에 동시에 결합할 수 있는 항체
하지만, 두 HC(Heavy Chain)와 두 LC(Light Chain)를 같이 발현시키면 총 10가지 이상의 조합이 가능하며, 바람직한 이중특이성 항체는 극히 일부.
-> 따라서 HC, LC 조합을 통제하는 엔지니어링 전략이 필요.
1️⃣ Knob-into-Hole (KiH) 전략
이중항체를 만들 때,
서로 다른 두 HC (heavy chain)가 정확하게 결합하도록 유도하기 위한 구조적 설계 방법
🖐 왜 이 전략이 필요한가?
IgG 항체는 두 개의 동일한 HC + 두 개의 동일한 LC로 구성됨
그런데 BsAb에서는 서로 다른 HC1과 HC2를 만들고 싶음
하지만 그냥 발현시키면
- HC1–HC1
- HC2–HC2
- HC1–HC2 (우리가 원하는 조합)
→ 이렇게 3가지 조합이 무작위로 섞임 → 순도 ↓, 정제 어려움
✅ 해결책: "Knob"과 "Hole" 도입
| 용어 | 설명 |
|---|---|
| Knob | 한쪽 HC의 CH3 도메인에 큰 아미노산 곁사슬을 돌출시켜 돌기처럼 만듦 |
| Hole | 다른쪽 HC에는 작은 아미노산으로 공간을 파서 홈을 만듦 |
→ 결과
HC1(knob) + HC2(hole) 조합만 잘 맞고,
HC1–HC1, HC2–HC2는 입체적으로 맞지 않아서 결합 안 됨
장점
- 간단한 돌연변이로 매우 안정적인 이질체 유도
- 단백질의 원래 Fc 기능(ADCC, CDC 등)도 대부분 보존 가능
단점
- LC mismatch는 방지 못함 → 별도의 light chain 제어 전략 필요
2️⃣ Interchain Disulfide Bond(이황화 결합) 추가
CH3 도메인 사이에 인공적인 이황화 결합(Cys–Cys)을 형성할 수 있도록 Cysteine 돌연변이를 삽입
-> 오직 이종 HC 간에만 이황화 결합이 형성되도록 설계
장점
- KiH보다 결합 안정성 높음 (구조적으로 고정)
- 열 안정성(Tm) 증가
단점
- Cys 돌연변이 도입 시 잘못된 이황화 결합 가능성 존재
- 산화/환원 조건에 민감
3️⃣ Electrostatic Steering (정전기 반발/유인)
같은 HC끼리는 반발, 다른 HC끼리는 흡인되도록 CH3 도메인 전하 재설계
4️⃣ cFAE (Controlled Fab-Arm Exchange)
서로 다른 두 항체의 팔만 교환하여, 이중특이성 항체(bispecific Ab)를 만드는 기술
🖐 IgG4의 자연 Fab-arm 교환
IgG4는 CH3 도메인 결합이 약하고, hinge 부위가 유연해서
체내에서 half-Ab 상태로 분리되었다가, 다른 IgG4와 Fab-arm을 교환함
-> 두 항원이 붙은 "functionally monovalent" 항체가 생성됨
🖐 IgG1 항체를 이용한 cFAE
IgG1은 원래 Fab-arm 교환을 하지 않음
하지만 실험실(in vitro)에서 특정 환원 조건 + CH3 변이를 주면
→ IgG1도 Fab-arm을 교환하게 만들 수 있음
이렇게 만든 게 cFAE 기반 이중항체(BsAb)
✅ 과정 요약
1️⃣ 두 parental IgG1 항체 (단일항원 타겟)을 준비
2️⃣ 환원 조건 (mild reducing condition)
→ 두 항체를 half-antibody (반쪽 항체) 상태로 만듦
→ heavy chain 2개가 분리됨
3️⃣ 교차 결합
→ 다른 항체의 반쪽과 섞어서 서로 다른 HC가 만나도록 함
4️⃣ 산화 조건으로 복원
→ 다시 결합시켜 이종 이량체(HC1-HC2)가 생기도록 함
5️⃣ CH3 도메인 변이 추가
→ 이종 heavy chain끼리만 잘 결합하도록 유도
(HC1–HC2는 안정 / HC1–HC1은 불안정)
장점
- Co-expression 필요 없음 → 두 항체 정제 후 섞기만 하면 됨
- Light chain mismatch 없음
→ Fc 기능 그대로 유지 + light chain은 자연 짝으로 유지 - 실제 임상용 BsAb 제작에 사용됨
단점
- in vitro 작업 필요 (세포 내 생산이 아님)
- 항체 구조에 따라 일부 항체는 잘 교환되지 않을 수 있음
5️⃣ Light Chain Control (LC 오조합 방지)
🖐 왜 Light Chain 문제가 생기는가?
이중항체를 만들 때,
보통 Heavy Chain(HC) 2종 + Light Chain(LC) 2종을 발현해야 함
→ 하지만 아무 HC와 아무 LC가 서로 아무렇게나 결합할 수 있음
→ 원하지 않는 조합 (mispaired Fab)이 많이 생김
1️⃣ κ–λ bodies
서로 다른 kappa/lamda light chain을 사용하여 구분 가능하게 만듦
🔎 ex)
하나의 Fab → kappa light chain (κ) 사용
다른 Fab → lambda light chain (λ) 사용각각의 light chain을 인식하는 항체로 순차적으로 정제 (serial affinity purification) 가능
🛑 But
κ와 λ가 모두 들어간 구조는 체내 안정성이나 발현 효율 문제가 생길 수 있음
2️⃣ Common LC (공통 경쇄 사용)
두 HC가 동일한 LC에 결합 가능하도록 설계 (LC는 하나만 사용)
🛑 But
두 항체가 같은 LC로 최적화되기 어려움
→ 항원 결합력이 떨어질 수 있음
→ 스크리닝으로 성능 좋은 공통 LC 찾아야 함
3️⃣ CrossMab 기술
Fab 내에서 LC 도메인을 교차(crossover)시켜, 잘못된 조합 방지
🔎 ex)
Fab 영역 내부에서 LC와 HC 도메인 일부를 서로 바꿈 (crossover)LC의 VL ↔ HC의 CL (또는 CH1 ↔ VL)
→ 구조적으로 HC1은 LC1만, HC2는 LC2만 결합 가능하게 만듦
구조적으로 HC와 올바른 LC가 짝지어지도록 유도
🛑 But
구조적 복잡성 ↑
일부 Fab 구조에서는 crossover가 기능을 저해할 수 있음
✅ Developability 기준
항체나 단백질이 의약품으로 개발될 수 있는지 평가하는 기준
- 구조적으로 안정한가?
- 대량 생산 가능한가?
- 체내에서 오래 살아남고, 면역반응 일으키지 않는가?
→ 아무리 항원에 잘 붙어도, developability가 낮으면 실제 치료제로 못 씀
1️⃣ Post-translational modification (PTM) 문제 적은 도메인
🖐 PTM이란?
단백질이 세포 내에서 만들어진 후에 가공되는 변화
🔎 ex)
글라이코실화(glycosylation), 산화(oxidation), 탈아민화(deamidation) 등
✅ PTM이 문제되는 이유
의도하지 않은 구조 변화
→ 효능 변화, 안정성 저하, 면역원성 유발 가능
✅ 개발 적합한 항체는?
PTM이 일어날 가능성이 적은 서열을 가짐
🔎 ex)
N-linked glycosylation motif (N-X-S/T)를 피함
CDR에 methionine이나 asparagine이 많으면 산화·탈아민화 위험 ↑
2️⃣ Solubility, Aggregation, Clipping 등의 위험 낮을 것
🖐 Solubility (용해도)
항체가 물에 잘 녹아야 체내에서도 잘 돌아다니고, 생산/정제도 쉬움
🖐 Aggregation (응집)
항체들이 서로 달라붙어 뭉치는 현상
면역반응 유도, 효능 감소, 안전성 문제 발생 가능
🖐 Clipping
단백질 일부가 효소에 의해 잘려나가는 현상
구조적 불안정 → 기능 상실
3️⃣ Neoepitope (새로운 항원 결정기) 없도록 설계 → ADA 유도 최소화
🖐 Neoepitope란?
단백질의 새로운 서열 구조나 표면이 면역계에 의해 '낯선' 것으로 인식되는 부위
특히 humanization하거나 CDR grafting할 때 잘 생김
B세포 or T세포가 이를 외부 단백질로 오해 →
→ ADA (Anti-Drug Antibody) 생성
→ 약효 감소, 알레르기 반응, 약물 중단
✅ 개발 적합한 항체는?
체내 자기 단백질처럼 보이도록 설계
Neoepitope가 생기지 않게 CDR, framework 조합 신중하게 설계